


Abstract
Multidrug-associated protein 2 (MRP2) is an efflux transporter that is expressed at the bile canalicular membrane. To allow in vitro to in vivo extrapolation of the contribution of MRP2 toward hepatic disposition of its substrates, data on the interindividual variability of hepatic MRP2 protein expression are required. Therefore, we quantified the expression of MRP2 in the University of Washington (UW) human liver bank (n = 51) using a modified version of a previously validated liquid chromatography/tandem mass spectrometry assay. An unlabeled (LTIIPQDPILFSGSLR) and stable isotope-labeled (LTIIPQDPILFSGSL[13C615N1]R) surrogate peptide for MRP2 were used as the calibrator and internal standard, respectively. After isolation of the membrane fraction from the liver tissue, in-solution tryptic digestion was conducted. Quality control samples created by spiking human serum albumin or pooled human liver (n = 51) matrix with three different MRP2 synthetic peptide concentrations generated error and precision values of less than 15%. As determined by the surrogate peptide, the average MRP2 expression in the UW liver bank samples was 1.54 ± 0.64 fmol/μg liver membrane protein and was found to be independent of age (7–63 years) or sex. A single nucleotide polymorphism in the promoter region (rs717620), previously thought to affect MRP2 expression, did not influence hepatic expression of MRP2. In contrast, the single nucleotide polymorphism 21214G>A (V417I; rs2273697) was associated with significantly higher hepatic MRP2 expression.
Introduction
Transporter-mediated biliary clearance (also termed as phase III drug elimination) is an important route of elimination for many xenobiotics, endobiotics, and their conjugated or unconjugated metabolites (Mizuno and Sugiyama, 2002; Giacomini et al., 2010). Factors affecting this pathway can have a significant impact on the hepatic concentration, clearance, and toxicity of drugs or their metabolites (Stieger et al., 2000; Kostrubsky et al., 2001; Endres et al., 2006, 2009; Feng et al., 2009; Ohtsuki et al., 2012). In humans, the measurement of the in vivo biliary clearance of drugs is difficult unless the intestine is catheterized (Bergman et al., 2010). Thus, in vitro methods such as sandwich-cultured human hepatocytes are often used to estimate the contribution of biliary clearance to the overall clearance of drugs. To improve such in vitro to in vivo extrapolation through physiologically based pharmacokinetic predictions (e.g., SimCYP), it is crucial to quantify the expression of transporters mediating the efflux of drugs and their metabolites. MRP2, encoded by the ABCC2 gene, is an important member of these efflux transporters, mediating the biliary efflux of a wide variety of drugs (e.g., fexofenadine, statins, spiramycin) and their phase II metabolites (glutathione and glucuronide conjugates) (Tian et al., 2007, 2008; Ieiri et al., 2009; Jemnitz et al., 2010). Although the expression of MRP2 in human livers has been previously quantified (Li et al., 2008, 2009; Ohtsuki et al., 2012), the small sample size did not provide an accurate estimate of the interindividual variability in the expression of this transporter. Here, we report the quantification of MRP2 expression in the University of Washington (UW) liver bank (51 samples) using liquid chromatography/tandem mass spectrometry (LC/MS/MS) methodology described previously (with few modifications) (Li et al., 2008, 2009). In addition, we determined whether the available information on genetic variation in ABCC2 in the liver bank was related to the level of MRP2 expression.
Materials and Methods
Chemicals and Reagents.
High-performance liquid chromatography-grade acetonitrile and other solvents were purchased from Thermo Fisher Scientific (Waltham, MA). Formic acid was purchased from Sigma-Aldrich (St. Louis, MO). The protein quantification BCA kit and the in-solution digestion kit were purchased from Thermo Fisher Scientific. The ProteoExtract Native Membrane Protein Extraction Kit and human serum albumin (HSA) were purchased from Calbiochem (Temecula, CA). A 16-mer synthetic peptide (LTIIPQDPILFSGSLR) representing a MRP2 tryptic peptide fragment and the stable isotope-labeled (SIL) internal standard (LTIIPQDPILFSGSL[13C615N1]R) were obtained from New England Peptides (Boston, MA).
Human Liver Samples.
Human liver samples [n = 51, age 7–63 years; 27 males and 24 females, all white, with the exception of one Asian male (HL165) and three non-Hispanic black males (HL104, HL105, and HL137)] of the Human Liver Bank of the University of Washington School of Pharmacy were investigated. Procurement, characteristics, and storage of these human livers have been described previously (Paine et al., 1997). At the time of collection, these livers were labeled as normal (22), fatty (18), fibrotic (5), with possible acute injury to the tissue (4), with iron deposition (HL141), or poorly perfused during the harvesting procedure (HL140) (Fig. 2A). Because of the anonymous nature of these samples, their use was classified as nonhuman subjects research by the UW Human Subjects Division.
Human Liver, Calibrator, and Quality Control Sample Preparation.
Isolation of the membrane fraction and subsequent tryptic digestion of MRP2 were conducted as described previously with the following minor modifications (Li et al., 2009). In brief, ∼100 mg of the liver tissue was homogenized in 2-ml extraction buffer I (ProteoExtract native membrane protein extraction kit; Calbiochem, Temecula, CA) in the presence of the protease inhibitor cocktail and incubated at 4°C for 10 min with gentle shaking. The homogenate was then centrifuged at 16,000g for 15 min at 4°C. The supernatant was removed, and the pellet was resuspended in the extraction buffer II of the kit with 10 μl of protease inhibitor cocktail, which was followed by incubation at 4°C with gentle shaking for 30 min. The suspension was centrifuged at 16,000g for 15 min at 4°C, and the supernatant containing membrane proteins was stored at −80°C until analysis.
Protein concentration in the isolated membrane fraction was determined by the BCA protein assay kit (Thermo Fisher Scientific). At the time of analysis, the membrane fraction was diluted to a working concentration of 2 μg/μl, and 20 μl of the extracted membrane fraction (equivalent to 40 μg of protein) was subsequently reduced with 10 mM dithiothreitol and alkylated with iodoacetamide in 50 mM ammonium bicarbonate digestion buffer. After the addition of the SIL internal standard, the protein samples were digested by trypsin in a final volume of 40 μl at 37°C for 4 h and then at 30°C for 14 h. At the end of digestion, the samples were acidified with equal amounts of 50% acetonitrile in H2O containing 0.1% formic acid (80-μl total sample volume) and centrifuged at 5000g for 20 min before LC/MS/MS analysis. The calibration curve was prepared by spiking HSA matrix (40 μg) with 20, 30, 40, 60, 80, 100, and 125 fmol of MRP2 peptide (in acetonitrile/water, 50:50) and subjected to tryptic digestion. Assay accuracy and precision were performed in triplicate at three different quality control (QC) concentrations of 25, 50, and 75 fmol of the peptide in 40 μg of HSA protein or pooled human liver membrane protein (n = 51) and then subjected to tryptic digestion. Bench-top stability of the MRP2 standard in the presence of HSA was measured at 6 h for 50 and 75 fmol/40 μg of protein. Likewise, autosampler stability was performed by reinjecting samples after 24-h storage at 6°C. Stability of MRP2 standard was investigated after three freeze-thaw cycles in HSA matrix.
LC/MS/MS Analysis.
The LC system consisted of a triple quadrupole mass spectrometer (API4000; Applied Biosystems, Foster City, CA) coupled to a Shimadzu LC (SLC-10A VP) system (Shimadzu, Wood Dale, IL) and an HTS PAL LEAP autosampler (LEAP Technologies, Carrboro, NC). The tryptic digest equivalent to 5 μg of membrane protein (10 μl) was injected onto the column (75 × 4.6 mm C8 column containing 5μ size beads, 100-Å pore size; Phenomenex, Torrance, CA) and eluted at 0.4 ml/min by a mobile phase with initial conditions of 92% A (water containing 0.1% v/v formic acid) and 8% B (acetonitrile containing 0.1% v/v formic acid) held for 4 min, followed by a linear gradient of 8% B to 27% B over the next 8 min and held until 15.1 min followed by wash and re-equilibration time. The parent-to-product transition for the MRP2 peptide monitored represents the doubly charged parent ion (LTIIPQDPILFSGSLR) 2H+ (m/z 886.1) to single charged product ions (m/z 1330.9 and 989.5) and a doubly charged ion (664.9). Likewise, the transition selected for the SIL internal standard peptide to the same doubly charged parent ion was m/z 889.5 to 1337.5, 996.6, and 668.5. The instrument settings of the API4000 triple quadruple mass spectrometer were as follows: ion spray voltage, 5.5 kV; temperature, 550°C; declustering potential, 56 V; collision energy, 43 V; entrance potential, 10 V; and collision cell exit potential, 36 V. Data were processed by integrating the appropriate peak areas generated from the reconstructed ion chromatograms for the 16-mer analyte peptide and the SIL internal standard peptide using the Analyst 1.4.1 software (Applied Biosystems, Foster City, CA).
Genotyping.
We determined whether variant alleles of MRP2 were associated with MRP2 expression. In brief, genomic DNA was extracted from individual livers. The samples were genotyped using Affymetrix DMET Plus Array (Santa Clara, CA) according to the manufacturer's protocol. Individual genotypes were resolved, and minor allele frequencies were determined using the Affymetrix DMET Plus console (version 1.1) using the Dynamic Genotype Boundaries algorithm (Y. Lin, K. Thummel, A. Rettie, and M. Rieder, unpublished results).
Results and Discussion
Assay Validation.
Among the three different transitions used for the MRP2 peptide standard (886.1 → 1330.9, 989.5 and 664.9) and MRP2 SIL internal standard (889.5 → 1337.5, 996.6 and 668.5), the transition 886.1 → 1330.9 for the MRP2 standard and 889.5 → 1337.5 for MRP2 SIL internal standard were most intense, consistently qualified QC parameters (see below) and, hence, were monitored during each experiment. The retention time for the MRP2 peptide standard and SIL internal standard was 14.1 min. The calibration curve prepared in HSA matrix showed a linear response over the range of the MRP2 peptide concentration used (0.5–3.125 fmol/μg protein).
The lower limit of quantification, defined as the lowest concentration of spiked MRP2 peptide in matrix with error and precision less than or equal to 20%, was 0.5 fmol/μg protein. Irrespective of the matrix used (pooled human liver membrane protein or HSA), the error and precision in the quantification of the QC samples were ≤15% (Fig. 1) at the three different concentrations tested. These data indicate the robustness of the assay and independence from the matrix used. Stability data (bench top, freeze-thaw, and autosampler) showed no degradation of the peptide (89.3–111.1% of the normal value).

Accuracy [% mean relative error (%RE) ± S.D. (A)], intraday (B), and interday (C) precision [% coefficient of variation (%CV)] of QC samples prepared in pooled human liver membrane protein (n = 51) or human serum albumin are shown. Data are mean of triplicate determinations. The listed amount represents the amount injected on-column in 5 μg of protein.
MRP2 Expression in the UW Liver Bank.
MRP2 expression was found to be an average of 1.54 ± 0.64 fmol/μg liver membrane protein (Fig. 2A). This is higher than the previous results in a smaller batch (n = 15) of human liver samples (0.6 ± 0.3 fmol/μg membrane protein) (Li et al., 2009) but similar to a recent report on 17 human livers (1.46 ± 0.65 fmol/μg liver membrane protein) (Ohtsuki et al., 2012). The larger sample size provides a better estimate of the interindividual variability in MRP2 expression. It is noteworthy that the interindividual variability in expression of MRP2 in the UW liver bank is modest (6.6-fold; 0.6–4.1 fmol/μg membrane protein). In fact, the majority of the liver samples (n = 36) had a narrower range of expression (1–2 fmol/μg membrane protein). Our yield of total membrane protein was 4.16 ± 0.77 mg/100 mg of liver tissue.
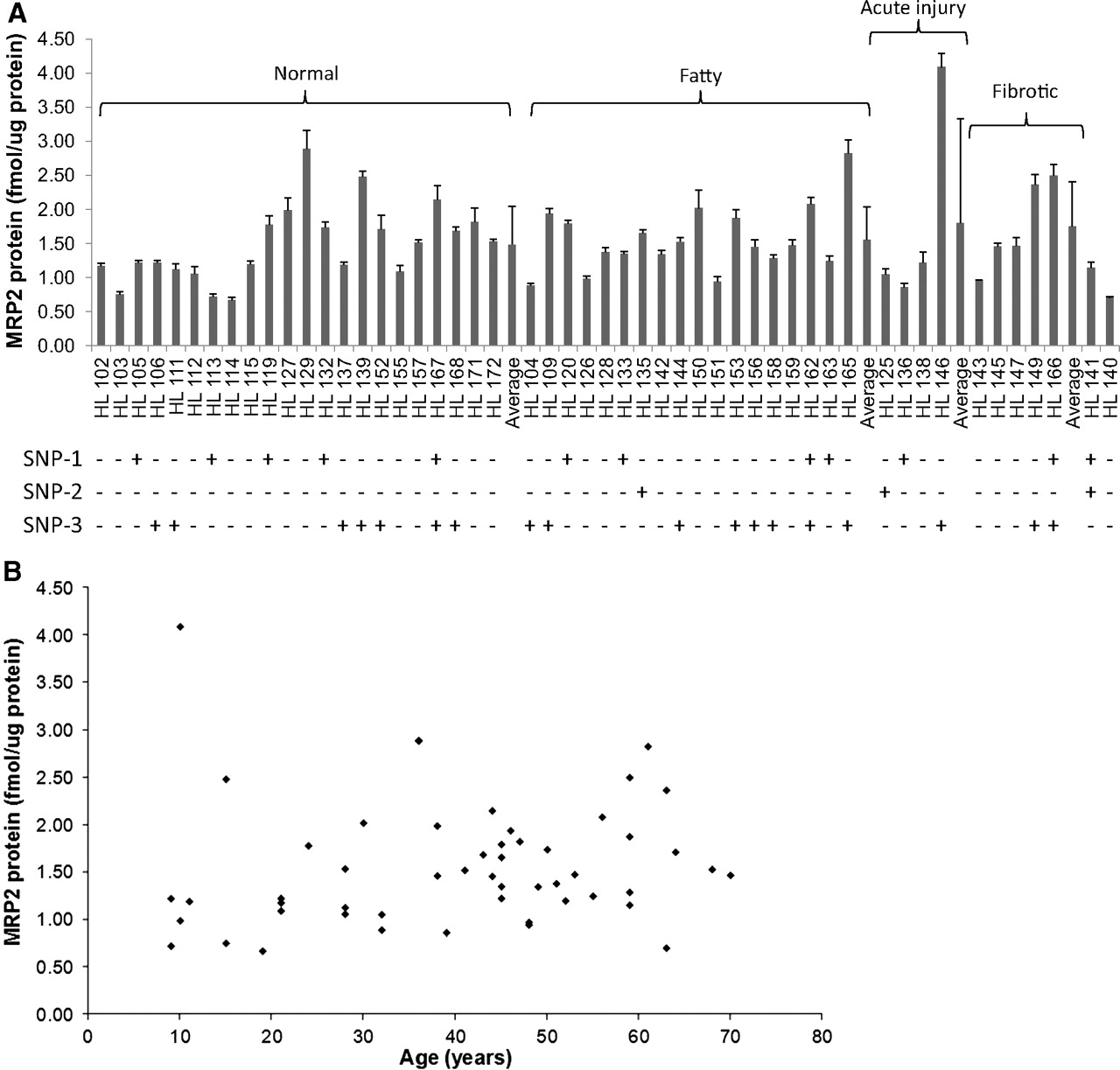
Expression of MRP2 in individual hepatic tissue from the UW human liver bank (n = 51 donors) categorized according to their condition. The average values for each category are also shown. SNP-1, SNP-2, and SNP-3 represent −24C>T (rs717620), 68693G>A(C1515Y; rs8187710) and 21214G>A(V417I; rs2273697), respectively. Expression of MRP2 in individual hepatic tissue (A) and its correlation with age (B) are shown. Data are mean ± S.D. of triplicate determinations.
Correlation of Subject Characteristics with Hepatic MRP2 Expression.
MRP2 expression (Fig. 2A) did not correlate with age (Fig. 2B) or sex. However, because our liver bank does not extend to children <7 years, we cannot conclude whether younger children may have a different level of MRP2 expression. There was no statistical difference observed in MRP2 expression in healthy (n = 22) versus fatty (n = 18) livers. It is noteworthy that HL146 (male, age 10 years) with acute injury and abnormal liver pathology (no additional information was available for this subject) showed the highest MRP2 expression (4.1 ± 0.2 fmol/μg liver membrane protein; Fig. 2A). Liver samples of donors with similar age or less than 10 years exhibited MRP2 expression in the adult range (Fig. 2B). The MRP2 expression in male livers (n = 27, 1.48 ± 0.72 fmol/μg membrane protein) did not differ significantly from those obtained from females (n = 24, 1.61 ± 0.56 fmol/μg membrane protein). There is insufficient ethnic variability in our liver bank to comment on the impact of ethnicity on MRP2 expression.
Correlation of MRP2 Polymorphism and MRP2 Protein Expression.
Genotyping revealed polymorphisms for ABCC2 at the following sites: 1846A>T(Y39F; rs927344), 21214G>A(V417I; rs2273697), −24C>T (rs717620), 36351T>G(L849R; rs45494393), 53395T>A(V1188E; rs17222723), 61606C>T(I1324I; rs3740066), 64260G>T(V1430V; rs1137968), 67932C>T(H1496H; rs8187707), and 68693G>A(C1515Y; rs8187710) in the promoter, coding and noncoding regions. Among these, two SNPs (i.e., −24C>T and 68693G>A) have been associated with decreased MRP2 activity, activity that was assessed indirectly (Wojnowski et al., 2005; Sookoian et al., 2009; Lubomirov et al., 2010; Franke et al., 2011). Here, we report the comparison of MRP2 expression in human livers harboring those SNPs reported to cause change in activity. We found that MRP2 expression in livers harboring −24C>T (11 heterozygotes, 1 homozygote) was not significantly different (1.55 ± 0.54 fmol/μg membrane protein) from the wild-type livers (1.56 ± 0.66 fmol/μg membrane protein), although Franke et al. (2011) found that this SNP resulted in increased erythromycin (a MRP2 substrate) metabolism as assessed by the breath test. The other SNP (68693G>A) was not present in high enough frequency to determine its impact on MRP2 expression. However, Meier et al. (2006) have shown that this SNP is associated with increased MRP2 expression. It is noteworthy that the SNP 21214G>A (V417I; rs2273697; 16 heterozygotes, 2 homozygotes), which has not been reported to affect MRP2 expression, showed a significantly (P < 0.004) higher expression of MRP2 (1.87 ± 0.77 fmol/μg membrane protein; n = 18) versus the wild-type livers (1.34 ± 0.38 fmol/μg membrane protein; n = 27). This SNP has been associated with greater central nervous system adverse effects (but not efficacy) of carbamazepine, presumably because of lower MRP2 activity (Kim et al., 2010). It is possible that this SNP increases the expression of MRP2 while reducing its activity. None of the livers harbored SNPs implicated in the Dubin-Johnson Syndrome (Suzuki and Sugiyama, 2002).
In summary, this is the first study to investigate MRP2 quantification in a large number of human livers that have been extensively phenotyped for cytochrome P450 activity and expression (Paine et al., 1997; Lin et al., 2002; Lalovic et al., 2004; Naraharisetti et al., 2010; Kaspera et al., 2011). Such MRP2 quantification data combined with MRP2 activity/expression data in cell lines expressing recombinant MRP2 or in hepatocytes will be a tremendous asset toward using physiologically based pharmacokinetic models to predict drug (or metabolite) hepatic exposure and clearance. To do so, it is important to have data on the expression of other transporters (e.g., solute carrier) and metabolizing enzymes (e.g., P450s) expressed in the liver bank. Such studies are in progress in our laboratory.
Authorship Contributions
Participated in research design: Deo, Prasad, Balogh, Lai, and Unadkat.
Conducted experiments: Deo and Prasad.
Performed data analysis: Deo, Prasad, and Unadkat.
Wrote or contributed to the writing of the manuscript: Deo, Prasad, Balogh, Lai, and Unadkat.
Acknowledgments
We thank Dr. Yvonne Lin for providing the genotyping data and Dale Whittington for assistance in the development of the LC/MS/MS assay.
Footnotes
This work was supported by the National Institutes of Health National Institute of General Medical Sciences [Grant GM32165]; a grant from Pfizer Inc.; and the Drug Metabolism, Transport and Pharmacogenomic Research program of the School of Pharmacy.
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
ABBREVIATIONS:
- MRP2
- multidrug resistance-associated protein 2
- UW
- University of Washington
- LS/MS/MS
- liquid chromatography/tandem mass spectrometry
- HSA
- human serum albumin
- QC
- quality control
- SNP
- single nucleotide polymorphism
- SIL
- stable isotope label.
- Received November 9, 2011.
- Accepted February 8, 2012.
- Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}