


Abstract
Morphine elimination involves UDP-glucuronosyltransferase (UGT) catalyzed conjugation with glucuronic acid to form morphine 3- and 6-glucuronides (M3G and M6G, respectively). It has been proposed that UGT2B7 is the major enzyme involved in these reactions, but there is evidence to suggest that other isoforms also catalyze morphine glucuronidation in man. Thus, we have characterized the selectivity and kinetics of M3G and M6G formation by recombinant human UGTs. UGT 1A1, 1A3, 1A6, 1A8, 1A9, 1A10, and 2B7 all catalyzed M3G formation, but only UGT2B7 formed M6G. The kinetics of M3G formation by the UGT1A family isoforms was consistent with a single enzyme Michaelis-Menten model, with apparent Km values ranging from 2.6 to 37.4 mM. In contrast, M3G and M6G formation by UGT2B7 exhibited atypical kinetics. The atypical kinetics may be described by a model with high- and low-affinity Km values (0.42 and 8.3 mM for M3G, and 0.97 and 7.4 mM for M6G) from fitting to a biphasic Michaelis-Menten model. However, a multisite model with an interaction between two identical binding sites in a negative cooperative manner provides a more realistic approach to modeling these data. According to this model, the respective binding affinities (Ks) for M3G and M6G were 1.76 and 1.41 mM, respectively. These data suggest that M6G formation may be used as a selective probe for UGT2B7 activity, and morphine glucuronidation by UGT2B7 appears to involve the simultaneous binding of two substrate molecules, highlighting the need for careful analysis of morphine glucuronidation kinetics in vitro.
Conjugation with glucuronic acid is an important metabolic pathway for the inactivation and elimination of a myriad of compounds, including drugs, dietary chemicals, environmental pollutants, and endobiotics. In particular, glucuronidation represents an important clearance mechanism for drugs from all therapeutic classes (Miners and Mackenzie, 1991). Glucuronidation reactions are catalyzed by enzymes of the UDP-glucuronosyltransferase (UGT1) gene superfamily. The individual forms of UGT (isoforms) tend to exhibit distinct, but overlapping, substrate specificities. Twenty-eight human UGT genes have been identified to date, and these have been classified into families and subfamilies based on evolutionary divergence (Mackenzie et al., 1997). However, only 14 of the known human UGTs appear to be catalytically active: UGT 1A1, 1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2A1, 2B4, 2B7, 2B15, 2B17, and 2B28.
Morphine is the preferred opioid for the relief of moderate to severe pain. Conversion to morphine 3- and 6-glucuronides (M3G and M6G, respectively) accounts for approximately two-thirds of the elimination of a parenteral dose of morphine in humans (Milne et al., 1996). Morphine 3-glucuronidation is the dominant pathway, and metabolic clearance to M3G is, on average, 5.4-fold higher than metabolic clearance to M6G. Although the liver appears to be the principal organ responsible for morphine glucuronidation in vivo, the gastrointestinal tract may contribute significantly to first-pass extraction after oral administration (Milne et al., 1996). Given the therapeutic importance of morphine and recognition that M6G also possesses analgesic activity, identification of the human UGT isoform(s) involved in the formation of the morphine glucuronides has attracted considerable interest. UGT2B7 has been shown to catalyze the conversion of morphine to both M3G and M6G and has been proposed as the major isoform involved in the glucuronidation of morphine in man (Coffman et al., 1997, 1998).
Although morphine is occasionally used as a substrate probe for UGT2B7, there have been no systematic studies of the UGT isoforms involved in morphine 3- and 6-glucuronidation. Indeed, there is evidence indicating that other isoforms have the capacity to glucuronidate morphine (Green et al., 1998; Cheng et al., 1999), and the apparent biphasic M3G formation kinetics in human liver microsomes (Miners et al., 1988) further suggests that multiple isoforms may catalyze the formation of this metabolite. We describe studies that aimed to characterize the selectivity and kinetics of M3G and M6G formation by recombinant human UGTs.
Materials and Methods
Chemicals. Morphine hemisulfate was a gift from Faulding Pharmaceuticals (Adelaide, Australia). M6G was purchased from Ultrafine Chemicals (Manchester, UK), and M3G, UDP-glucuronic acid (UDPGA), 4-methylumbelliferone (4MU), and 4MU-β-d-glucuronide (4MUG) were purchased from Sigma-Aldrich (St. Louis, MO). Other chemicals and reagents were of analytical reagent grade.
Expression of UGTs. UGT 1A1, 1A3, 1A6, 1A8, 1A9, 1A10, 2B4, 2B7, 2B10, and 2B15 were stably expressed in HK293 cells. Cells were transfected with cDNAs cloned into the pEF-IRES-puro6 expression vector, and microsomes from cells expressing UGT1A family isoforms were prepared as described previously (Sorich et al., 2002). However, the microsome preparation procedure resulted in significant loss of activity of UGT2B family isoforms. Thus, cells expressing UGT 2B4, 2B7, 2B10, and 2B15 were lysed by probe sonication, 3 × 1 s with cooling on ice for 3 min between pulses, and lysates were stored at -70°C in phosphate buffer (0.1 M, pH 7.4) until measurement of enzyme activity.
Measurement of M3G and M6G Formation. Incubations contained morphine (6–11 concentrations; see Fig. 1), UDPGA (10 mM), MgCl2 (5 mM), and microsomal or lysate protein (0.45 mg) in phosphate buffer (0.1 M, pH 7.4) in a total volume of 0.3 ml. Reactions were initiated by the addition of UDPGA and performed at 37°C for 90 min. Following the addition of perchloric acid (11.6 M, 0.012 ml) and 4MUG (the assay internal standard, 1 pmol), samples were vortex mixed and then centrifuged (1500g for 10 min). A 0.2-ml aliquot of the supernatant fraction was added to an Eppendorf tube containing KOH (2 M, 0.012 ml). M3G and M6G concentrations were quantitated using a specific high-performance liquid chromatographic method with fluorescence detection (Stone et al., 1998). Overall assay imprecision was <10% for substrate concentrations in the range 0.025 to 10 mM.

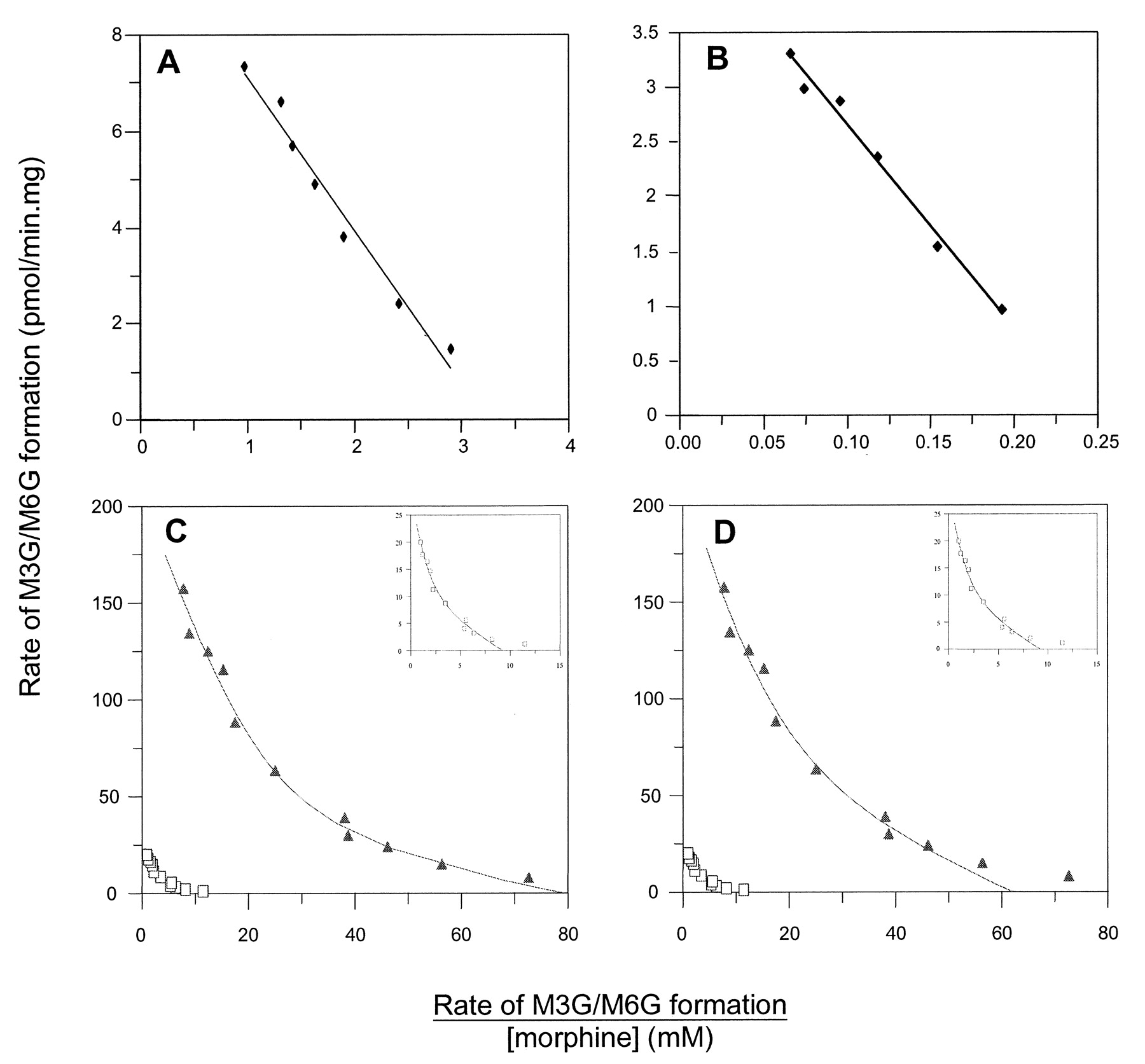
Eadie-Hofstee plots for M3G formation by UGT1A1 (A) and UGT1A3 (B); panels C and D show Eadie-Hofstee plots for M3G (▴) and M6G (□) formation by UGT2B7 from fitting to a biphasic Michaelis-Menten (C) and two-site (D) model.
The inset in panels C and D are a magnification of the plots for M6G formation. Points represent experimental data, and curves were generated from model-fitting.
Measurement of 4MUG Formation. To confirm expression of individual UGTs, lysates or microsomes prepared from HK293 cells were tested for glucuronidation of the nonselective UGT substrate 4MU using a previously published fluorescence assay (Sorich et al., 2002). The 4MU concentrations used were 100 and 1000 μM. Overall assay imprecision was <3% at these substrate concentrations.
Data Analysis. All data points represent the mean of duplicate
estimations. The kinetic parameters Km and
Vmax were calculated from untransformed data by nonlinear
least-squares regression using GraFit (Erithacus Software, Horley, Surrey,
UK). Data were fitted to the Michaelis-Menten equations for a single- and
two-enzyme model with a weighting of 1/y, and to the two-site model
described by Kenworthy et al.
(2001).
 where [S] is substrate concentration, Ks is the substrate
dissociation constant and α and β are binding factors that reflect
changes in Ks and product formation
(Kp), respectively. Goodness of fit was determined by
comparison of statistical parameters (χ2 and Akaike information
criterion values) between the models and a reduction on the standard errors of
the parameter estimates. Kinetic data are reported in
Table 1 as mean ± S.E.
of fit.
where [S] is substrate concentration, Ks is the substrate
dissociation constant and α and β are binding factors that reflect
changes in Ks and product formation
(Kp), respectively. Goodness of fit was determined by
comparison of statistical parameters (χ2 and Akaike information
criterion values) between the models and a reduction on the standard errors of
the parameter estimates. Kinetic data are reported in
Table 1 as mean ± S.E.
of fit.
Kinetic constants for morphine 3- and 6-glucuronide formation by human recombinant UDP-glucuronosyltransferases calculated from Michaelis-Menten models
Results
Apart from UGT2B4 and UGT2B10, 4MU glucuronidation activity was measurable (>10 pmol/min · mg at a 4MU concentration of 100 or 1000 μM) for all recombinant UGT isoforms (data not shown). Formation of 4MUG by UGT2B4, but not UGT2B10, was however confirmed using a more sensitive radiometric thin-layer chromatographic procedure (Jin et al., 1997). UGT 1A1, 1A3, 1A6, 1A8, 1A9, 1A10, and 2B7 all catalyzed morphine 3-glucuronidation, whereas UGT 2B4, 2B10, and 2B15 lacked the capacity to glucuronidate morphine. M3G formation kinetics by the UGT1A family isoforms were consistent with a single-enzyme Michaelis-Menten model, with apparent Km values ranging from 2.6 to 37.4 mM. Derived kinetic constants are summarized in Table 1, and Eadie-Hofstee plots for representative low- (UGT1A1) and high- (UGT1A3) affinity isoforms are shown in Fig. 1 (panels A and B). Considerable variation was observed in Vmax values (Table 1). However, relative expression of the isoforms, which presents difficulties with UGT, was not determined, and the significance of this observation remains to be assessed.
In contrast to the UGT1A family isoforms, M3G formation by UGT2B7 (in cell lysates) exhibited atypical kinetics (Fig. 1, C and D). (M3G formation kinetics by microsomes prepared from HK293 cells expressing UGT2B7 also exhibited nonhyperbolic kinetics, but the lower activity of the microsomal preparation precluded full kinetic characterization.) Derived kinetic constants from fitting to a biphasic Michaelis-Menten model are given in Table 1. Apparent Km values for the high- and low-affinity components were 0.42 and 8.3 mM, respectively. Vmax values for the high- and low-affinity components differed 7.8-fold. Derived parameters using the two-site model were: Ks, 1.76 ± 0.78 mM; Vmax, 109 ± 36 pmol/min · mg; α, 3.40 ± 1.07; and β, 1.97 ± 0.50. UGT2B7 was the only isoform that mediated morphine 6-glucuronidation. M6G formation also exhibited atypical kinetics (Fig. 1, C and D). Derived Km values from the biphasic Michaelis-Menten model fit (Fig. 1C) were 0.97 ± 0.29 mM and 13.7 ± 4.0 mM (Table 1). Parameters generated from the two-site model (Fig. 1D) were: Ks, 1.41 ± 0.56 mM; Vmax, 13.9 ± 3.8 pmol/min · mg; α, 4.19 ± 1.15; and β, 2.00 ± 0.42.
Discussion
Morphine has been shown previously to be a substrate for UGT2B7, and the formation of M3G has been used as an index of human liver microsomal UGT2B7 activity. However, data presented here suggest that formation of M6G, rather than M3G, is a selective probe for UGT2B7 activity in human tissues since multiple isoforms potentially contribute to morphine 3-glucuronidation. Formation of M3G by UGT1A3 and 1A8 is consistent with previously published data (Green et al., 1998; Cheng et al., 1999). A number of human UGTs were not screened for their capacity to glucuronidate morphine in this study, namely, UGT 1A4, 2A1, 2B11, 2B17, and 2B28. UGT1A4 has been reported previously not to metabolize morphine (Green et al., 1998), and like UGT2B10, UGT2B11 is an “orphan” enzyme apparently lacking catalytic activity (Jin et al., 1993; Beaulieu et al., 1998). UGT2A1 is expressed predominantly in olfactory epithelium (Tukey and Strassburg, 2000) and thus would not contribute significantly to morphine elimination in vivo. Roles for UGT2B17 and 2B28 in drug metabolism have not been established but cannot be discounted.
Of the isoforms capable of metabolizing morphine, UGT 1A1, 1A3, 1A6, 1A9, and 2B7 are expressed in the liver and may potentially contribute to hepatic clearance. However, the low affinities of UGT 1A1, 1A6, and 1A9 suggest that involvement of these isoforms is likely to be minor at best. Expression of UGT 1A8 and 1A10 is limited to the gastrointestinal tract, and UGT 1A1, 1A3, 1A6, 1A9, and 2B7 have also been identified in this tissue (Tukey and Strassburg, 2000). Data reported here are consistent with a contribution of the gastrointestinal tract to the first-pass extraction of morphine (Milne et al., 1996).
The kinetics of M3G and M6G formation by UGT2B7 were adequately described by either a biphasic Michaelis-Menten model or a two-site model based on the simultaneous binding of two substrate molecules at identical sites. Both approaches have been adopted previously to analyze the biphasic kinetic behavior of certain reactions catalyzed by recombinant cytochromes P450 (Houston and Kenworthy, 2000; Hutzler and Tracy, 2002). The values for α derived for the two-site model were both >1, indicating the decreased binding affinity of a second substrate molecule in the presence of the first at the active site (negative cooperativity) (Houston and Kenworthy, 2000). The equivalence of the two binding sites is indicated by similar binding affinities (Ks) of M3G and M6G for the active site. The factor β reflects changes in the effective catalytic rate constant (Kp) as the result of an interaction between the two occupied sites. Whenever the equivalence of two substrate binding sites is assumed, the value of β is 2 (Vmax is equivalent to 2Kp[E]t, where [E]t is the total enzyme concentration), which was observed for both M3G and M6G formation. Although atypical kinetics have been described for drug metabolism by recombinant cytochromes P450 (Shou et al., 2001; Galetin et al., 2002; Hutzler and Tracy, 2002), the phenomenon of negative cooperativity has been rarely observed for either substrate kinetics or the effect of a modifier. Previous studies of morphine glucuronidation by UGT2B7 have not alluded to biphasic kinetics. Apparent Km values reported previously for M3G and M6G formation ranged from 0.47 to 1.0 mM and 0.55 to 1.3 mM, respectively (Coffman et al., 1997, 1998).
This report provides confirmation of atypical kinetic properties of human UGTs. Using UGT2B7, evidence of negative cooperativity is presented, consistent with multisite interactions between morphine and this enzyme. The two-site model provides more realistic insight to the interactions of morphine at the UGT2B7 active site than the biphasic Michaelis-Menten model, although both generate reasonable fit. Studies with UGTs require careful design and detailed experimentation to delineate kinetic properties.
Footnotes
-
↵1 Abbreviations used are: UGT, UDP-glucuronosyltransferase; UDPGA, UDP-glucuronic acid; M3G, morphine 3-glucuronide; M6G, morphine 6-glucuronide; 4MU, 4-methylumbelliferone; 4MUG, 4MU-β-d-glucuronide.
-
This work was supported by a grant from the National Health and Medical Research Council of Australia. A.N.S. was the recipient of an Australian Postgraduate Award (Industry), funded by the Commonwealth of Australia Department of Employment, Education, Training and Youth Affairs in collaboration with Faulding Pharmaceuticals.
- Received February 26, 2003.
- Accepted May 21, 2003.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}