


Abstract
An analysis of reported hepatic abundances of CYP3A4 and 3A5 indicated that values determined by immunoquantification using commercially available, unpurified recombinant enzymes as standards are significantly lower than those determined using purified enzymes or human liver microsomes characterized with lysosomal peptides (CYP3A4: mean 45 versus 121 pmol/mg protein, p < 0.01; CYP3A5: mean 28 versus 83 pmol/mg protein, p < 0.05). When immunoquantifying cytochromes P450 (P450s), it is assumed that the holoprotein (holo)/apoprotein ratio is the same in the samples and the standard. Estimates of holo/apoprotein ratios from data reported for a range of P450s purified from human liver and non-commercial recombinant systems indicated less than complete and variable heme coupling dependent on enzyme and system.
The absolute abundances of cytochrome P450 (P450) enzymes in human liver, expressed as amount of enzyme per milligram of microsomal protein, are required for scaling of in vitro data on drug metabolism by recombinant human P450 systems (rhP450) to in vivo hepatic clearance (Barter et al., 2007). The use of intersystem extrapolation factors (Proctor et al., 2004) allows the differences in intrinsic activity (per unit P450) between rhP450 and human liver enzymes to be accounted for. The corrected rhP450 in vitro data must then be combined with the abundance of the appropriate P450 enzyme in human liver as part of the scaling process. If such abundances are established for large numbers of individual human livers, it is possible to combine this information with activity per unit enzyme obtained with rhP450 systems to predict the distribution of drug clearance across populations, without the need to assess enzyme activity directly in large numbers of liver samples (Rostami-Hodjegan and Tucker, 2007). The accuracy of such in vitro-in vivo predictions will clearly depend on the fidelity of estimates of individual P450 abundances in human liver. The latter are usually determined by immunoblotting (Laemmli, 1970) or enzyme-linked immunosorbant assay. More recently, mass spectrometric methods have also been proposed for P450 quantification in human samples (Lane et al., 2004; Alterman, 2005; Jenkins et al., 2006). However, all of these methods measure apoprotein, which comprises active protein in which heme is incorporated (holoprotein), and that in which it is not. By contrast, only holoprotein is measured by carbon monoxide (Omura and Sato, 1964) or dithionite difference spectroscopy (Matsubara et al., 1976). A variety of protein standards have been used to immunoquantify P450s in human liver microsomes, ranging from enzyme purified to electrophoretic homogeneity from either liver microsomes or rhP450s (Guengerich and Turvy, 1991; Shimada et al., 1994), human liver microsomal standard (HLMSTD) (Westlind Johnsson et al., 2003) characterized using lysozyme-peptide conjugates (Edwards et al., 1998), to (more recently) commercially available (unpurified) rhP450 systems (King et al., 2003; Galetin et al., 2004; Wang et al., 2005). An assumption in using any of these standards is that the holoprotein/apoprotein ratio is the same in the samples and the standard. To our knowledge the implications of this assumption have never been assessed.
The aims of this study were 2-fold: first, to carry out a meta-analysis of CYP3A4 and 3A5 abundance values determined using different calibration standards, and second, to assess holoprotein/apoprotein ratios from studies reporting the purification of P450s from human liver and recombinant systems.
Materials and Methods
Abundances of CYP3A4 and CYP3A5 in Human Liver. Values of human hepatic CYP3A4 and CYP3A5 abundance were collated from two electronic databases, MEDLINE (http://www.nlm.nih.gov/databases/databases_medline.html) and Web of Knowledge (http://wok.mimas.ac.uk/), and personal files of the authors (1990-2006) containing references from “Current Contents” and “Reference Updates”. The authors of the original articles were contacted directly when further information was required. Only data from adult Caucasians (>16 years) were included, and sources were verified to exclude duplication of individual data in the analysis. Geometric mean values of abundance were used to represent central tendency since the frequency distributions of the data were not normal (Kolmogorov-Smirnov test; SPSS v12, SPSS Inc., Chicago, IL). Overall weighted mean (WX̄) values of CYP3A4 and CYP3A5 abundance were calculated using eq. 1: 
where there are J sources of data, n samples in each source, and x̄ is the mean value from each data source. The weighted geometric mean values (WX̄geo) were calculated using eq. 2: 
where CV is the coefficient of variation (%).
Heterogeneity in the data was assessed from the homogeneity number (HMG), calculated using eqs. 3 to 5: 


where wj is the weight of each study based on the variance of the data and VWX̄ is the variance of the weighted mean of all observations (1-J).
The significance of differences between CYP3A4 and CYP3A5 abundance values determined from studies using rhP450 standards and those using HLMSTD or purified enzyme were assessed by Student's t test (Data Analysis Toolpack, Microsoft Office Excel 2003; Microsoft, Redmond, WA).
Determination of Holoprotein/Apoprotein Ratios of Purified P450s. The MEDLINE database was searched for reports of P450 enzyme purification from both human liver microsomes and recombinant expression systems. The molecular mass (kDa) of each P450 (CYP1A2, 58.3; 2A6, 56.5; 2B6, 56.3; 2C8, 55.8; 2C9, 55.6; 2C19, 56.0; 2D6, 55.8; 2E1, 56.9; 3A4, 57.3; 3A5, 57.1) was used to calculate the expected specific enzyme content, assuming 100% holoprotein. The actual percentage holoprotein content of each preparation was calculated using eq. 6 by comparing the measured value of the specific P450 content per milligram of total protein determined by spectroscopy with the expected value 
Deviation of holoprotein protein contents from 100% were assessed for each P450 using the z-test. Differences in holoprotein content between P450s were assessed by one-way analysis of variance followed by Tukey's b post hoc test.
Results
The analysis of CYP3A4 abundance values was based on 384 livers from 13 separate studies (Table 1). The overall weighted geometric mean value was 82 pmol/mg microsomal protein, and there was a 10-fold difference between mean estimates from different studies. The homogeneity test gave an HMG of 37 (p < 0.001), indicating that the reported average values of abundance in these studies did not conform to a unimodal distribution. Accordingly, the mean value of CYP3A4 abundance determined from studies using rhP450 systems as the calibration standard was significantly lower (p < 0.01) than the mean value from studies using characterized human liver microsomes or purified enzyme (45 versus 121 pmol/mg microsomal protein). In all of the studies included in the meta-analysis, rhP450 enzymes were obtained from commercial sources.
Literature values of mean CYP3A4 abundance
The analysis of CYP3A5 abundance values was based on 45 livers from seven separate studies (Table 2). The overall weighted geometric mean value was 55 pmol/mg microsomal protein, and there was an 8-fold difference between mean estimates from different studies. An HMG value of 53 indicated significant (p < 0.001) heterogeneity in the results of the different studies. Accordingly, the mean value of CYP3A5 abundance determined from studies using rhP450 systems as the calibration standard was significantly lower (p < 0.05) than the mean value from studies using characterized human liver microsomes or purified enzyme (28 versus 83 pmol/mg microsomal protein).
Literature values of mean CYP3A5 abundance
The percentage contributions of holoprotein to total P450 protein purified from human liver were found to be significantly less than 100% (p < 0.01 for CYP2C19 and p < 0.001 for CYP2C8, 2C9, 2D6, and 3A4) and the mean holoprotein/apoprotein ratio for CYP2D6 preparations was significantly (p < 0.05) less than that of the other P450 preparations (Fig. 1A). There were also indications of intersubject differences in holoprotein proportion, as exemplified by the analysis of data for CYP3A4 in preparations from three different livers purified in the same laboratory (Fig. 1B). The percentage contributions of holoprotein to total P450 protein purified from rhP450 systems are shown in Fig. 2. Unlike the rhP450 preparations used in the meta-analysis of CYP3A4 and CYP3A5 abundance, rhP450s used to obtain purified enzyme were from noncommercial sources.
Discussion
The study of CYP3A4 and CYP3A5 abundances in human liver indicated that the use of different protein standards may result in different values, with commercial rhP450 standards providing generally lower estimates than characterized human liver microsomes and purified enzyme preparations (Tables 1 and 2). A possible explanation for the latter observation is that different standards have different holoprotein/apoprotein ratios. Thus, if the standard contains a lower proportion of holoprotein than that in the samples, the same immunoblot signal will indicate a lower amount of active enzyme, resulting in an underprediction of active P450 abundance in the samples.

A, mean percentage contribution of P450 enzyme holoprotein to total P450 protein in preparations purified from human liver tissue. Standard deviations are shown by error bars where the number of livers was sufficient for calculation. B, interindividual variability in percentage contribution of holoprotein to total CYP3A4 protein purified from three individual livers (from Guengerich et al., 1986). (References to the data for each of the P450s in A are 1A2: Distlerath et al. 1985; 2A6: Yun et al., 1991; 2C8: Wang et al., 1980; Wrighton et al., 1987b; Lasker et al., 1998; 2C9: Shimada et al., 1986; Kawano et al., 1987; Lasker et al., 1987, 1998; Komori et al., 1988; Sandhu et al., 1993; 2C19: Lasker et al., 1987, 1998; 2D6: Gut et al., 1984, 1986; Dislerath et al., 1985; 2E1: Lasker et al., 1987; Wrighton et al., 1987a; 3A4: Wang et al., 1983; Watkins et al., 1985; Guengerich et al., 1986; Kawano et al., 1987; Komori et al., 1988; Lin et al., 2002.)
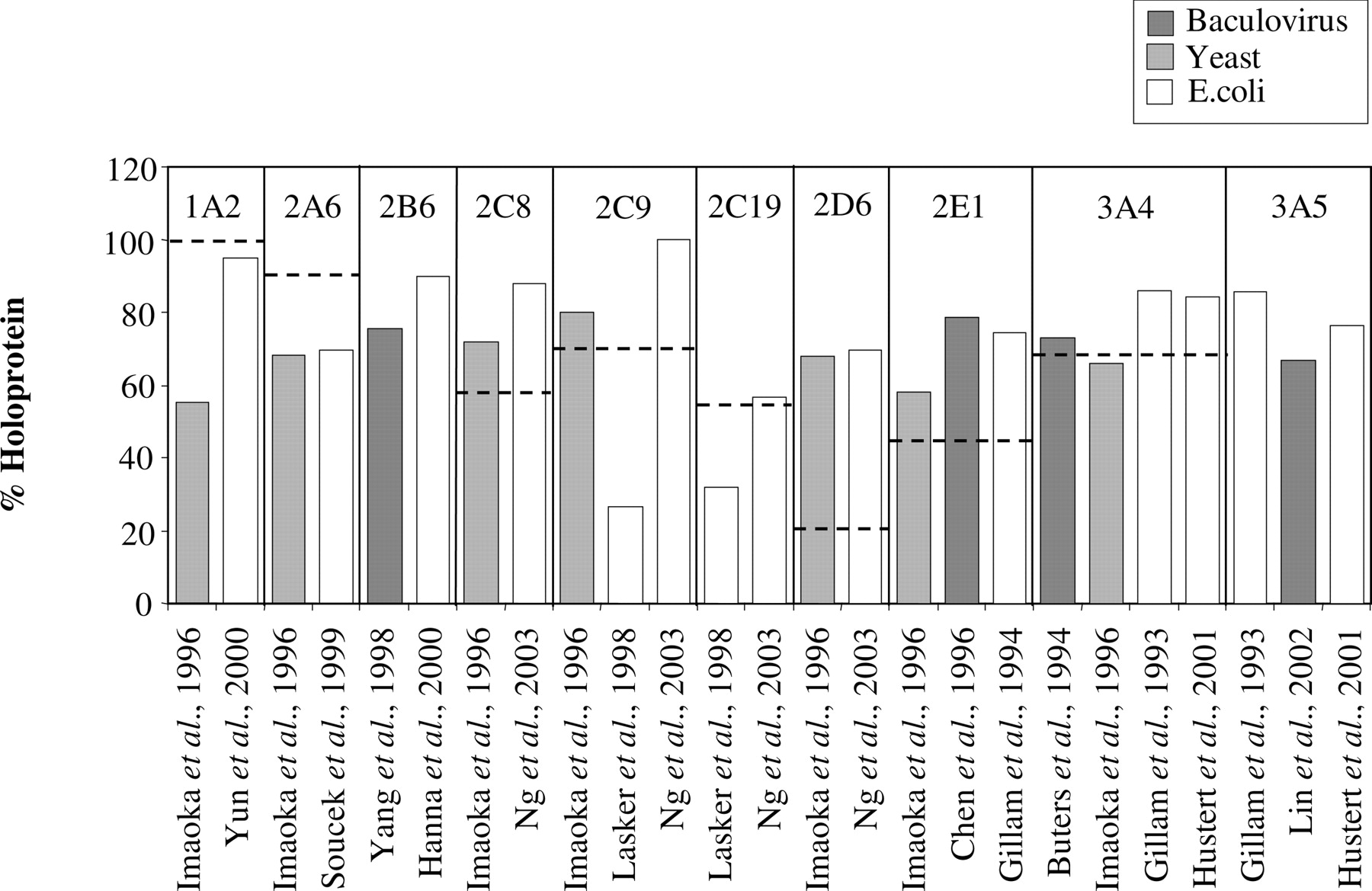
Percentage holoprotein contribution in preparations purified from recombinant P450 expression systems. For comparison, the dotted lines indicate the mean percentage holoprotein contribution observed for each P450 enzyme when purified from human liver tissue.
Estimates of the holoprotein/apoprotein ratios of purified P450s from human liver indicate incomplete heme coupling, and differences in this respect between specific P450s and possibly between individual livers (Fig. 1). Although incomplete protein purification may explain the findings, it is unlikely that contamination would be more than 10% since loading of 5 μg of protein on a gel is usually sufficient to detect bands from other proteins running separately from the “pure” enzyme. Thus, incomplete purification cannot account for the greater than 10% differences in holoprotein/apoprotein ratios observed in many cases. If incomplete purification can be discounted, the observations are either due to incomplete heme incorporation in vivo or experimental artifact (uncoupling of the heme from P450 protein during the purification process), or both. The latter assumes that the uncoupling happens to different extents in different purified systems. In any event, the findings have important implications for the immunoquantification of P450 abundances in human liver samples if there is a mismatch of the holoprotein/apoprotein ratio in standards and samples. Depending on the standard used, there could be either under- or overprediction of P450 abundance.
The estimated holoprotein contents of purified rhP450s also suggest incomplete heme coupling. The majority of purified rhP450 preparations have a holoprotein/apoprotein ratio similar to that seen in the human liver preparations, suggesting that these would be suitable to use as standards for immunoquantification (Fig. 2). However, the CYP2D6 preparations had higher holoprotein contents than that seen in the human liver preparations. Therefore, if these were used as a standard, a significant overprediction in CYP2D6 abundance would result. It should be noted that these rhP450 standards have not been produced for commercial use. The low estimations of P450 abundance indicated in Tables 1 and 2 suggest that commercially available recombinantly expressed P450 systems would have a lower holoprotein content than those shown in Fig. 2.
The observation that estimates of CYP3A abundances are lower when using commercially available rhP450 systems as standards relative to human liver standards could reflect a lower holoprotein/apoprotein ratio in these systems. To establish whether this is the case, it would be necessary to show that the commercial rhP450 systems give a greater immunoblot signal for the same level of spectrally determined holoprotein compared with purified liver enzyme. Preliminary studies have suggested that this is the case for CYP3A4 and CYP3A5 rhP450 systems (Wilson et al., 2005; Perrett et al., 2006). This work is ongoing, and we hope that the outcome will enable determination of appropriate correction factors to apply when measuring enzyme abundance with rhP450 systems. The utility of such factors would also depend on the extent of variability in the holoprotein/apoprotein ratio between individual liver samples. It is possible that the ratio of holoprotein to apoprotein might also be affected by genotype since single residue changes can markedly affect protein stability.
Acknowledgments
We thank Dr. K. Rowland Yeo for assistance with the meta-analysis of P450 abundances.
Footnotes
-
H.F.P. and Z.E.B. were supported by Simcyp Ltd., Sheffield, UK, and EU Framework 6 (BIOSIM).
-
doi:10.1124/dmd.107.015743.
-
ABBREVIATIONS: P450, cytochrome P450; rhP450, recombinant human cytochrome P450; HLMSTD, human liver microsomal standard; WX̄, weighted mean; HMG, homogeneity number; VWX̄, variance of the weighted mean.
- Received March 9, 2007.
- Accepted June 25, 2007.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}