


Abstract
Bevirimat [BVM, PA-457, 3-O-(3′,3′-dimethylsuccinyl)-betulinic acid], a new anti-human immunodeficiency virus drug candidate, is metabolized to two monoglucuronides [mono-BVMG (I) and mono-BVMG (II)] and one diglucuronide (di-BVMG) both in vivo and in vitro. UDP-glucuronosyltransferase (UGT) reaction screening, enzyme kinetics, and species differences for the glucuronidation of BVM in vitro were investigated with pooled human liver microsomes (HLMs) and human intestinal microsomes (HIMs), animal liver microsomes, and 12 recombinant human UGT isoforms. Glucuronidation of BVM with HLMs predominantly involved the formation of mono-BVMG (I) (Vmax = 61 pmol/min/mg protein, Km = 27 μM) and mono-BVMG (II) (Vmax = 48 pmol/min/mg protein, Km = 16 μM). Di-BVMG was also observed but was a minor metabolite. HIMs mainly revealed glucuronidation to form mono-BVMG (II) (Vmax = 90 pmol/min/mg of protein, Km = 8.3 μM). UGT1A3 predominantly formed mono-BVMG (I) (Vmax = 65 pmol/min/mg of protein, Km = 13 μM), whereas UGT1A4 is a less active isoform (Vmax = 1.8 pmol/min/mg of protein, Km = 5.6 μM). UGT2B7 was involved in the formation of both mono-BVMG (I) (Vmax = 6.1 pmol/min/mg of protein, Km = 6.0 μM) and mono-BVMG (II) (Vmax = 6.5 pmol/min/mg of protein, Km = 7.8 μM). Among the animal liver microsomes examined, all species (rat, mouse, dog, and marmoset) demonstrated conjugation to form both mono-BVMG (I) and mono-BVMG (II), with dog liver microsomes exhibiting a higher formation rate for mono-BVMG (I), whereas marmoset liver microsomes showed a higher formation rate for mono-BVMG (II). The data suggest a primary role of UGT1A3 for the glucuronidation of BVM.
Glucuronidation, a conjugation reaction catalyzed by UDP-glucuronosyltransferases (UGTs), is one of the major phase II drug metabolic pathways that contribute to drug biotransformation (Radominska-Pandya et al., 1999; King et al., 2000; Ritter, 2000; Tukey and Strassburg, 2000; Fisher et al., 2001). Acyl (ester) glucuronidation is an important metabolic pathway of carboxylic acid-containing endogenous compounds, such as bilirubin and retinoic acid, and drugs, such as nonsteroidal anti-inflammatory drugs (NSAIDs), fibrate hypolipidemic agents (gemfibrozil and clofibric acid), diuretics (furosemide), anticonvulsants (valproate), and antibiotics (moxifloxacine and gemfloxacine) (Spahn-Langguth and Benet, 1992; Sallustio et al., 2000; Bailey and Dickinson, 2003; Shipkova et al., 2003; Sakaguchi et al., 2004). Identification of human UGTs responsible for glucuronidation of existing and novel drugs provides critical information about potential drug-drug interactions and possible genotypic varieties in drug disposition.
Bevirimat [BVM, PA-457, 3-O-(3′,3′-dimethylsuccinyl)-betulinic acid], a derivative of aliphatic triterpenic acid with two carboxyl groups (Fig. 1), represents a new class of antiretroviral drugs, termed maturation inhibitors, that block HIV maturation by inhibiting the final step in the processing of the HIV Gag protein (Li et al., 2003). Preclinical studies have shown that BVM retains full activity against viral strains resistant to currently available treatments and is effective in an animal model of HIV infection. BVM has demonstrated positive results in a phase I/II clinical trial in HIV-infected patients. After oral administration to rats, BVM is transformed into its acyl glucuronides (BVMGs) including two monoglucuronides [mono-BVMG (I) and mono-BVMG (II)] and one diglucuronide (di-BVMG) (Wen et al., 2006a). In vitro metabolism studies suggest little oxidative metabolism of bevirimat in human tissues (data not shown). Structural characterization by liquid chromatography-electrospray ionization-mass spectrometry (LC-ESI-MS) indicated that mono-BVMG (I) was glucuronidated at the C-28 carboxyl of BVM, whereas mono-BVMG (II) was conjugated at the dimethylsuccinic side chain of the C-3 position of BVM (Wen et al., 2006a). The objectives of this study were 1) to screen UGT enzyme responsibilities for the in vitro acyl glucuronidation of BVM using human liver microsomes (HLMs), human intestinal microsomes (HIMs), and 12 commercially available recombinant human UGT isoforms, 2) to estimate enzyme kinetics for the formation of acyl glucuronides of BVM catalyzed by the responsible recombinant human UGT isoforms, HLMs, and HIMs, and 3) to investigate species differences for the in vitro acyl glucuronidation of BVM in human and animal liver microsomes.

Chemical structures of BVM, BVM glucuronides, and internal standard (DSD). MW, molecular weight.
Materials and Methods
Chemicals. BVM and DSD [3-O-(3′,3′-dimethylsuccinyl)-dihydrobetulinic acid, a reduced analog of BVM] (Fig. 1), were provided by Panacos Pharmaceuticals (Gaithersburg, MD). DSD was used as the internal standard for the quantification of BVM and BVMGs. Uridine 5′-diphosphoglucuronic acid (UDPGA), magnesium chloride, d-saccharic acid 1,4-lactone (d-SL) (a specific β-glucuronidase inhibitor), phenylmethylsulfonyl fluoride (PMSF) (a carboxylesterase inhibitor), alamethicin, trifluoroacetic acid (TFA); bilirubin, etoposide, imipramine, diclofenac, zidovudine (AZT), trifluoperazine (TFP), 7-hydroxy-4-trifluoromethylcoumarin (7-HFC), 4-trifluoromethyl-7-hydroxycoumarin glucuronide (7-HFCG), and Tris-HCl, pH 7.4, were purchased from Sigma-Aldrich (St. Louis, MO). Pooled HLMs were obtained from In Vitro Technologies (Baltimore, MD). Pooled rat (RLMs), mouse (MLMs), and dog liver microsomes (DLMs) and 12 recombinant human UGTs (BD Supersomes enzymes: UGT1A1, -1A3, -1A4, -1A6, -1A7, -1A8, -1A9, -1A10, -2B4, -2B7, -2B15, and -2B17) were purchased from BD Gentest (Woburn, MA). Pooled HIMs were purchased from XenoTech (Lenexa, KS). Pooled marmoset liver microsomes (MMLMs) and pooled rat intestinal microsomes (RIMs) were prepared as described previously (Soars et al., 2001; Tallman et al., 2005). The protein contents were used as described in the data sheets provided by the manufacturers, except for MMLMs and RIMs, which were measured by a bicinchoninic acid protein assay kit from Pierce Chemical (Rockford, IL) using bovine serum albumin as a standard. Acetonitrile (ACN; HPLC grade) and methanol (MeOH; HPLC grade) were obtained from Mallinckrodt (Phillipsburg, NJ). Bond Elut C18 cartridges (100 mg, 1 ml) for solid-phase extraction (SPE) were obtained from Varian (Palo Alto, CA). All other chemicals and reagents were of analytical grade.
Assays for Enzyme Activities of Recombinant Human UGTs, HLMs, and HIMs. Enzyme activities of recombinant human UGTs (except UGT1A4), HLMs, and HIMs were measured by a HPLC-UV assay, using 7-HFC as a substrate according to the manufacturer's protocol with modifications. In brief, an incubation mixture (final volume 125 μl) containing 0.25 mg of protein/ml microsomes, 0.1 M Tris-HCl buffer, pH 7.4, and 25 μg/mg of protein alamethicin in the final incubation was preincubated on ice for 10 min. After the addition of MgCl2 (5 mM), d-SL (5 mM), and 7-HFC (100 μM), the incubation mixture was preincubated at 37°C for 5 min. The stock solution of 7-HFC was prepared in MeOH and then diluted with water. The final concentration of MeOH in the incubation was 1% (v/v). The reaction was initiated by the addition of UDPGA (5 mM) and incubated at 37°C in a shaking water bath for 30 min; then 100-μl aliquots of the incubation were withdrawn. The reaction was terminated, and microsomal protein was precipitated by the addition of 100 μl of ice-cold ACN-glacial acetic acid (HAc) [96:4 (v/v)]. After the removal of the protein by centrifugation at 15,000g for 10 min at 4°C, 50-μl aliquots of the supernatants were injected onto a HPLC column (Axxiom ODS, 150 × 4.6 mm i.d., 5 μm; Thomson, Clear Brook, VA) and separated at ambient temperature using a gradient elution with an Agilent HP 1100 LC system (Agilent Technologies, Palo Alto, CA). Elution was carried out by the following gradient program with solvent A (0.1% HAc) and solvent B (MeOH): 0 to 11 min, 10% B to 90% B; 11 to 12 min, 90% B; 12 to 13 min, 90% B to 10% B; and 13 to 15 min, 10% B. The flow rate was 1 ml/min, and the detection wavelength was 325 nm. The retention times of 7-HFCG and 7-HFC were 8.3 and 11.5 min, respectively. Quantification of the glucuronide product was performed by a comparison to the absorbance of an external standard curve for 7-HFCG.
The activity of recombinant human UGT1A4 was determined using TFP as a substrate. TFP was dissolved in water and its final concentration in the incubation was 200 μM. The incubation and HPLC instrumental conditions were the same as above, except the mobile phase solvents with solvent A (0.1% TFA) and solvent B (ACN containing 0.1% TFA) and detection at 256 nm. The retention times of TFPG and TFP were 8.8 and 9.6 min, respectively. Quantification of the glucuronide product was performed by comparing to the absorbance of an external standard curve for TFP, assuming that TFPG has an absorbance response similar to that of TFP under the experimental conditions.
Glucuronidation of BVM. In vitro glucuronidation of BVM was conducted in an incubation volume of 125 μl. Incubation conditions were initially optimized using HLMs for linear product formation with respect to substrate concentrations (0–100 μM), microsomal protein concentrations (0.25, 0.5, 1, and 2 mg/ml), incubation time (0.5, 1, 2, and 4 h), and activation of microsomes by alamethicin (10, 25, 50, and 100 μg/mg of protein) or detergent Brij 35 (0.1, 0.25, 0.5 and 1 mg/mg of protein). Preliminary experiments indicated that the glucuronide formation of BVM was linear up to 2 h of incubation time and 2 mg/ml microsomal protein. Both alamethicin and Brij 35 activated the glucuronidation of BVM. However, treatment with detergent Brij 35 exhibited a bell-shaped curve of activity, whereas alamethicin showed a linear increase of glucuronide formation up to 50 μg/mg of protein of alamethicin.
In brief, an incubation mixture containing 1 mg of protein/ml of microsomes, 0.1 M Tris-HCl buffer, pH 7.4, and 25 μg/mg of protein alamethicin in the final incubation was preincubated on ice for 10 min. After the addition of MgCl2 (5 mM), d-SL (5 mM), and substrate (BVM), the incubation mixture was preincubated at 37°C for 5 min. BVM stock solution (di-N-methylglucamine salt of BVM) was directly prepared with water. The reaction was initiated by the addition of UDPGA (5 mM) and incubated at 37°C in a shaking water bath for 60 min; then 50-μl aliquots of the incubation were withdrawn. The reaction was immediately terminated, and microsomal protein was precipitated by the addition of 200 μl of ice-cold ACN containing 0.5% formic acid (pH of the final incubation mixture was adjusted to 3 at this step to stabilize acyl glucuronides) and internal standard DSD (100 ng). After the removal of protein by centrifugation at 15,000g for 10 min at 4°C, the supernatants were transferred. After the addition of 1 ml of 0.1% HAc to reduce the organic solvent strength, the resultant supernatants were loaded onto a SPE cartridge preconditioned by washing with 1 ml of MeOH and then 1 ml of 0.1% HAc. The SPE cartridge was then washed with 1 ml of ACN-0.1% HAc [5:95 (v/v)] and subsequently eluted with 2 ml of ACN-0.1% HAc [90:10 (v/v)]. The eluant was dried with a stream of nitrogen at 45°C in a water bath, and the residue was reconstituted in 100 μl of ACN-0.1% HAc [80:20 (v/v)] for LC-ESI-MS assay. Preliminary experiments indicated that the reconstituted samples containing BVM, BVMGs, and internal standard (DSD) were stable at least for 72 h at 4°C.
Analysis of BVM and Its Glucuronides. Identification and quantification of BVM and BVMGs were performed as described previously (Wen et al., 2006a). In brief, HPLC separation was carried out using an Agilent HP 1050 LC system (Agilent Technologies) with a guard column (Zorbax RX-C8, 12.5 × 4.6 mm i.d., 5 μm; Agilent Technologies) and an analytical column (Zorbax RX-C8, 150 × 2.1 mm i.d., 5 μm; Agilent Technologies). HPLC conditions were as follows: mobile phase A, 0.1% formic acid-50 mM ammonium formate (pH ∼4.5) and B, ACN containing 0.1% formic acid; gradient elution, 0 to 10 min, 20% B to 90% B; 10 to 16 min, 90% B; and 16 to 18 min, 90% B to 20% B. The column temperature was ambient, the flow rate was 0.3 ml/min, and the injection volume was 10 μl. MS analysis was performed with an API 100 liquid chromatography/mass spectrometry system (PerkinElmer Sciex, Toronto, ON, Canada) with a TurboIonspray interface in the negative ESI ionization mode. Quantification of BVM and BVMGs was carried out by selective ion monitoring with [M-H]– for di-BVMG (m/z 935.6), mono-BVMGs (m/z 759.6), BVM (m/z 583.5), and internal standard DSD (m/z 585.5), respectively. Because no BVNG authentic standards were currently available, the ESI-MS responses and related quantitative coefficients of BVMGs were estimated by comparing the initial and released contents of BVM in rat bile (mainly containing BVMGs) with and without β-glucuronidase hydrolysis. The contents of BVMGs were then calculated using the calibration curves of BVM with the quantitative coefficients estimated, assuming that all forms of BVMGs have similar ESI-MS response ratios derived from the peak area ratio to that of internal standard (Wen et al., 2006a).
UGT Reaction Screening of BVM. UGT reaction screening of BVM was performed with 12 commercially available recombinant human UGTs, insect cell control (negative control) microsomes, HLMs, and HIMs, using a constant amount of microsomal protein (1 mg/ml) and two different concentrations of BVM (10 and 50 μM, respectively). Incubation conditions were the same as described earlier. Incubations without UDPGA or without substrate were also carried out. The incubation samples were extracted and purified by SPE as described above and then analyzed by LC-ESI-MS.
Enzyme Kinetics for the Glucuronidation of BVM. Apparent enzyme kinetic parameters (Km and Vmax) for the glucuronidation of BVM were determined with HLMs, HIMs, and recombinant human UGT1A3, -1A4, and -2B7, using various concentrations of BVM (0–100 μM) at fixed concentrations of 1 mg of protein/ml of microsomes, MgCl2 (5 mM), alamethicin (25 μg/mg of protein), d-SL (5 mM), and UDPGA (5 mM) in a 125-μl final incubation. Incubation conditions were similar to those for the glucuronidation of BVM described previously. After incubation, internal standard DSD (100 ng) was added to the samples. The samples were extracted and purified by SPE as described above, and the concentrations of BVM glucuronides were quantified by LC-ESI-MS. Apparent Km and Vmax were estimated by fitting the duplicate experimental data to a Michaelis-Menten equation: V = (Vmax × S)/(Km + S), where Km is the Michaelis-Menten constant, Vmax is the maximum velocity, and S is the substrate concentration, using a nonlinear least-squares regression method.
Species Differences for the Glucuronidation of BVM. Species differences for the in vitro glucuronidation of BVM were determined with HLMs, RLMs, MLMs, DLMs, and MMLMs, using a constant amount of microsomal protein (1 mg/ml) and two different concentrations of BVM (10 and 50 μM, respectively). HIMs were also studied. Incubation conditions were the same as described above. Incubations without UDPGA or without substrate were also performed. The incubation samples were extracted and purified by SPE as described earlier and then analyzed by LC-ESI-MS.
In Vitro Stability of BVMGs in UGT Reaction Media with HIMs and RIMs. Rat bile samples containing BVMGs (Wen et al., 2006a) were incubated in the glucuronidation reaction media (alamethicin, 25 μg/mg of protein; MgCl2, 5 mM; without UDPGA) with HIMs and RIMs (1 mg of protein/ml) at pH 7.4 (0.1 M Tris-HCl) and 37°C for 0, 0.5, 1, 2, and 4 h. Aliquots of the incubations with HIMs and RIMs were withdrawn at different time points and were prepared using the procedures of analysis for BVM and BVMGs. The contents of BVMGs and BVM in the incubations with and without inhibitors (d-SL and PMSF) were analyzed by LC-ESI-MS.
Inhibitory Effects of Typical Probe Substrates on BVM UGT Activity in HLMs. Six typical UGT probe substrates were investigated for their potential inhibitory effects on BVM UGT activity. Bilirubin (Bosma et al., 1994) and etoposide (Watanabe et al., 2003; Wen et al., 2006b) for UGT1A1, imipramine (Green et al., 1995, 1998; Nakajima et al., 2002) for UGT1A3 and -1A4, TFP (Green and Tephly, 1996; Dehal et al., 2001) for UGT1A4, AZT (Barbier et al., 2000; Court et al., 2003) for UGT2B7, and diclofenac (King et al., 2001; Sakaguchi et al., 2004) for UGT1A3 and UGT2B7 were used as the potential inhibitors. Bilirubin and etoposide stock solutions were prepared in DMSO, and the final concentrations of DMSO in the incubation was 1% (pre-experiments indicated that 1% of DMSO in the final incubation has no influence on the glucuronidation of BVM). Imipramine, TFP, AZT, and diclofenac stock solutions were prepared in water. The formation rates of BVM glucuronides in HLMs (1 mg of protein/ml) at a fixed concentration of BVM (25 μM) with and without inhibitors (0–500 μM) were determined. Incubation conditions were the same as those described previously for the glucuronidation of BVM. The 50% inhibitory concentration (IC50) was estimated graphically.
Results
Reaction Phenotyping of BVM Glucuronidation. Enzyme activities of 12 commercially available recombinant human UGTs, HLMs, and HIMs measured by HPLC-UV assays indicated that the UGT enzymes used in this study were active as per measurement of 7-HFC and TFP (for UGT1A4) is adequate (data not shown). Incubation of BVM (10 and 50 μM, respectively) with recombinant human UGTs, HLMs, and HIMs demonstrated that three UGT isoforms (UGT1A3, -1A4, and -2B7), HLMs, and HIMs catalyzed the acyl glucuronidation of BVM (Figs. 2 and 3). No glucuronidation of BVM was observed in the presence of the other nine UGT isoforms tested, insect cell control microsomes, and incubations without UDPGA (Fig. 3). UGT1A3, -1A4, and -2B7 catalyzed the formation of mono-BVMG (I), and this metabolite was formed at a much faster rate by UGT1A3 (∼60 pmol/min/mg of protein at 50 μM BVM) than the formation rates catalyzed by UGT1A4 and -2B7. The rate of mono-BVMG (I) formation in incubations containing HLMs (∼50 pmol/min/mg of protein at 50 μM BVM) was similar to that observed for UGT1A3. Under these conditions, only UGT2B7 seemed to catalyze the formation of both mono-BVMG (I) and mono-BVMG (II). A small amount of di-BVMG could be detected in incubations containing HLMs (results not shown), but it was not quantitatively detectable under the experimental conditions used in this study, presumably because of the low rate of BVM glucuronidation catalyzed by this microsomal sample. It is notable that both mono-BVMG (I) and mono-BVMG (II) were detected in approximately the same amount (based on peak areas) in incubations containing HLMs, whereas mono-BVMG (II) was the predominant product in incubations with HIMs (Figs. 2 and 3).

Representative LC-ESI-MS chromatograms for the formation of BVM glucuronides after incubation of 10 μM BVM with HLMs, HIMs, recombinant human UGTs (UGT1A3, 1A4, and 2B7), and insect control microsomes. SIM, selective ion monitoring.

Enzyme screening of acyl glucuronidation of BVM with HLMs, HIMs, recombinant human UGTs, and insect control microsomes. The concentrations of BVM in the final incubations were 10 and 50 μM, respectively. The formation rates of BVM glucuronides were determined as described under Materials and Methods. Data are expressed as the means of individual duplicate incubations in picomoles per minute per milligram of protein.
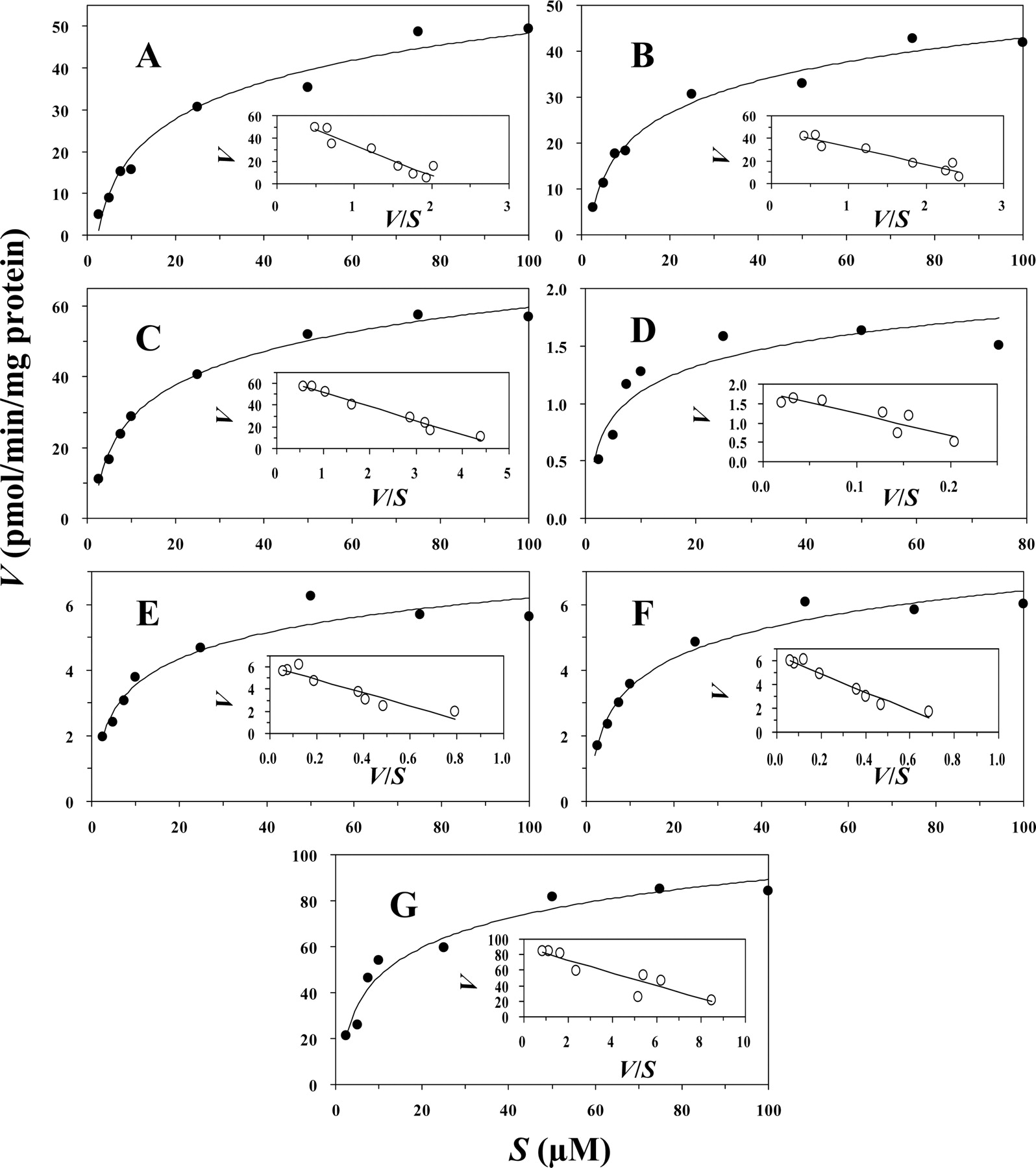
Enzyme Kinetic Parameters. The enzyme kinetic parameters for the glucuronidation of BVM were evaluated with microsomes containing recombinant human UGT1A3, -1A4, and -2B7, as well as with HLMs and HIMs (Fig. 4; Table 1). The apparent Vmax values for the formation of mono-BVMG (I) with UGT1A3 and HLMs were very similar (65 and 61 pmol/min/mg of protein, respectively). In contrast, the Vmax values for the same reaction catalyzed by UGT1A4 and -2B7 were at least 10-fold lower (1.8 and 6.1 pmol/min/mg of protein, respectively). However, the apparent Km values for the formation of mono-BVMG (I) by UGT1A3, -1A4, and -2B7 were very similar (13, 5.6, and 6.0 μM, respectively), whereas the Km value for the same reaction catalyzed by HLMs was 2- to 5-fold higher (27 μM); the greater Km in HLMs could be due to nonspecific binding differences caused by differing protein concentrations used in recombinant enzyme systems and liver microsomes. Mono-BVMG (I) was not quantitatively formed in the presence of HIMs under these conditions. The Vmax values for the formation of mono-BVMG (II) by UGT2B7, HLMs, and HIMs were quite different (6.5, 48, and 90 pmol/min/mg of protein, respectively); however, the Km values were similar (7.8, 16, and 8.3 μM, respectively). The formation of mono-BVMG (II) was not catalyzed by UGT1A3 or -1A4 in this study.

Enzyme kinetics of acyl glucuronidation of BVM in recombinant human UGTs (UGT1A3, -1A4, and -2B7), HLMs, and HIMs. The formation rates of BVM glucuronides were determined as described under Materials and Methods. Data are expressed as Data are expressed as the means of individual duplicate incubations in picomoles per minute per milligram of protein. A, mono-BVMG (I) for HLMs; B, mono-BVMG (II) for HLMs; C, mono-BVMG (I) for UGT1A3; D, mono-BVMG (I) for UGT1A4; E, mono-BVMG (I) for UGT2B7; F, mono-BVMG (II) for UGT2B7; G, mono-BVMG (II) for HIMs.
Kinetic parameters for the formation of acyl glucuronides of BVM with recombinant human UGTs and pooled human liver and intestinal microsomes
Data were measured by LC-ESI-MS and are expressed as the means of duplicates.
Species Differences. Liver microsomes isolated from five mammalian species, as well as HLMs and HIMs, were incubated with BVM (10 and 50 μM, respectively) to investigate the potential species-related differences in the formation of BVMGs observed in the UGT reaction phenotyping experiments (Table 2). The formation of di-BVMG was observed only in the incubations containing RLMs or DLMs, with much lower velocities than those of mono-BVMGs. The mono-BVMG (I) metabolite was formed at quite different formation rates in the incubations of the five species tested. DLMs catalyzed the formation of mono-BVMG (I) at the highest rates (86 and 140 pmol/min/mg of protein at 10 and 50 μM BVM, respectively). The rates of formation of mono-BVMG (I) catalyzed by HLMs, RLMs, and MLMs were relatively modest (12–50 pmol/min/mg of protein), whereas HIMs and MMLMs demonstrated very weak catalyzing activities (∼3 pmol/min/mg of protein). The mono-BVMG (II) metabolite was formed in the incubations for all of the microsomal samples used in this study. HIMs exhibited relatively higher rates (54 and 82 pmol/min/mg of protein at 10 and 50 μM BVM, respectively) in comparison with other species tested, whereas HLMs, RLMs, and MMLMs demonstrated moderate catalyzing activities (14–63 pmol/min/mg of protein). The rates of formation of mono-BVMG (II) catalyzed by MLMs (5.2 and 9.3 pmol/min/mg of protein) and DLMs (2.8 and 29 pmol/min/mg of protein) were relatively low.
Acyl glucuronide formation of BVM (10 and 50 μM, respectively) with different microsomal sources
Data were measured by LC-ESI-MS and are expressed as the means of duplicates.
In Vitro Stability of BVMGs in Incubation Media with Intestinal Microsomes. We have investigated the in vitro stability of acyl (ester) glucuronides of BVM (BVMGs) in glucuronidation reaction media with HLMs, RLMs, and MLMs, as well as in human, rat, and mouse plasma (Wen et al., 2006a), and reported that BVMGs are stable in the incubation media with liver microsomes and plasma in the presence of d-SL (a specific β-glucuronidase inhibitor) and/or PMSF (a carboxylesterase inhibitor). Because HLMs catalyze the glucuronidation of BVM to both mono-BVMG (I) and mono-BVMG (II), whereas HIMs catalyze mainly the glucuronidation of BVM to mono-BVMG (II), this led us to investigate the stability of BVMGs in intestinal microsomes. In vitro stability of BVMGs in glucuronidation reaction media with HIMs and RIMs demonstrated that, in the presence of d-SL and/or PMSF, BVMGs exhibited relatively high stability in incubation media with HIMs (Fig. 5, A, C, and E) and RIMs (Fig. 5, B, D, and F). In the absence of the inhibitors, when spiked into the inhibition media, all forms of BVMGs in HIMs (Fig. 5G) and two BVM monoglucuronides in RIMs (Fig. 5H) did not show a significant decline in comparison with their initial concentrations. Di-BVMG was labile to hydrolysis, particularly in RIMs. However, the absolute concentration level of di-BVMG was much lower than those of the two BVM monoglucuronides (data not shown). In addition, no significant interconversion between mono-BVMG (I) and mono-BVMG (II) was observed both in HIMs and RIMs in vitro. Moreover, little intramolecular acyl migration was detectable in vitro when assessed by susceptibility to β-glucuronidase cleavage (Wen et al., 2006a). Because the concentration level of di-BVMG in the rat bile tested was very low, small but significant increases in BVM resulting from the degradation of di-BVMG were found in this study (data not shown). These results are consistent with the previous in vitro stability data in human and animal liver microsomes and plasma (Wen et al., 2006a), suggesting that the monoglucuronides of BVM exhibit high stability in vitro. However, di-BVMG, the minor metabolite of BVM, is a bit more labile in liver and intestinal microsomes.
Inhibition of BVM Glucuronidation by UGT Probe Substrates. The inhibitory effects of bilirubin and etoposide (UGT1A1), imipramine (UGT1A3 and -1A4), TFP (UGT1A4), AZT (UGT2B7), and diclofenac (UGT2B7 and -1A3) on BVM UGT activity with HLMs are shown in Fig. 6. The estimated IC50 values for each substrate against both BVM monoglucuronides are summarized in Table 3. The results demonstrated that at 500 μM, bilirubin, etoposide, and AZT did not inhibit the formation of either BVM monoglucuronide. Imipramine and diclofenac inhibited the formation of both BVM monoglucuronides to a moderate degree (40–70%) when the concentrations of inhibitors were high, and diclofenac inhibited the formation of mono-BVMG (I) more than the formation of mono-BVMG (II) (70% versus 40%). TFP (500 μM) more effectively inhibited the formation of both BVM monoglucuronides to the same degree (>90%).
Estimated IC50 (μM) of typical inhibitors on BVM (25 μM) UGT activity in human liver microsomes
Discussion
Acyl glucuronides are the conjugation products of compounds containing a carboxylic acid functional group with the sugar, d-glucuronic acid. Human UGT1A1, -1A3, -1A6, -1A9, and -2B7 have been reported as the important catalysts involved in acyl glucuronidation of carboxylic acid-containing compounds, such as bilirubin, retinoic acid, NSAIDs, and others (Ritter, 2000; Sallustio et al., 2000; Shipkova et al., 2003; Sakaguchi et al., 2004). However, other UGT isoforms such as UGT1A7, -1A8, and -1A10, which are mainly expressed in extrahepatic tissues, should not be excluded. In this study, we first characterized the direct glucuronidation of BVM, a maturation inhibitor with two carboxylic acids, by human liver and intestinal microsomes and recombinant human UGTs (Figs. 2 and 3). HLMs catalyzed the formation of mono-BVMG (I) and mono-BVMG (II) at very similar rates, whereas HIMs catalyzed the rapid formation of mono-BVMG (II) and the relatively slow formation of mono-BVMG (I). Di-BVMG was formed only in the incubations containing HIMs, but it was not quantitatively detectable. UGT reaction phenotyping with 12 commercially available recombinant human UGTs, demonstrated that UGT1A3, -1A4, and -2B7, major human hepatic UGT isoforms, catalyzed the formation of mono-BVMG (I) and that this metabolite was formed more quickly by UGT1A3 than by UGT1A4 or -2B7. Although the enzyme contents of recombinant UGT1A3 and HLMs were not exactly known, their catalyzing activities were very similar. It is noteworthy that HIMs predominantly catalyzed the glucuronidation to form mono-BVMG (II). However, if recombinant UGT2B7, also present in human intestine, is capable of forming both BVM mono-glucuronides in vitro and is the only UGT isoform screened that produced mono-BVMG (II), it is not clear why HIMs primarily catalyzed the formation of mono-BVMG (II). The kinetic analysis for the formation of the two BVM monoglucuronides by human fractions and recombinant UGTs (Table 1) suggests that the formation of mono-BVMG (I) is catalyzed by a moderate-affinity, high-capacity enzyme present in HLMs. In addition, mono-BVMG (I) is also formed by UGT1A4 and -2B7, both of which seem to be moderate-affinity, low-capacity catalysts. The kinetics of mono-BVMG (II) formation seems to be quite different from those of mono-BVMG (I). The former was detected only in the presence of UGT2B7, HLMs, and HIMs. UGT2B7 also acted as a moderate-affinity, low-capacity catalyst (e.g., Km = 7.8 μM and Vmax = 6.5 pmol/min/mg of protein). HLMs behaved as a low-affinity catalyst of moderate capacity (e.g., Km = 16 μM and Vmax = 48 pmol/min/mg of protein) and HIMs also behaved as a low-affinity, moderate-capacity catalyst (e.g., Km = 8.3 μM and Vmax = 90 pmol/min/mg of protein), athough higher than HLMs.

In vitro stability of BVM glucuronides in UGT reaction media with HIMs and RIMs. Aliquots of rat bile samples containing BVM glucuronides were incubated in glucuronidation reaction media (without UDPGA) with HIMs and RIMs (1 mg of protein/ml) at 37°C for 0, 0.5, 1, 2 and 4 h. The concentrations of BVM and BVM glucuronides in the incubations were determined as described under Materials and Methods. Data (means of individual duplicate incubations) are expressed as peak area ratios relative to internal standard (DSD) in comparison with the controls at time 0. A (HIMs) and B (RIMs), with d-SL (5 mM) and PMSF (0.5 mM); C (HIMs) and D (RIMs), with d-SL (5 mM); E (HIMs) and F (RIMs), with PMSF (0.5 mM); G (HIMs) and H (RIMs), without d-SL and PMSF.
We also investigated the species differences for the in vitro acyl glucuronidation of BVM (Table 2). With respect to the selective formation and catalytic rates of the two BVM monoglucuronides, the incubations with RLMs yielded results that most closely resembled those obtained with HLMs. Formation of BVM diglucuronide in vitro may not be relevant to in vivo formation because of the active efflux of BVM monoglucuronides, as rat bile had very low levels of BVM diglucuronide (Wen et al., 2006a). DLMs had the highest formation rates, but favored mono-BVMG (I). Furthermore, the results obtained with MMLMs most closely resembled those obtained by HIMs, with mono-BVMG (II) being the preferred product.
Our previous experiments demonstrated that acyl glucuronides of BVM have relatively high in vitro stability in glucuronidation reaction media with HIMs and RIMs, as well as human and animal liver microsomes and plasma (Wen et al., 2006a). The relatively high stability of BVM glucuronides (Fig. 5) is possibly due to their structural characteristics (Fig. 1) with fully substituted α-carbons to the carboxyl groups resulting in minimal acyl migration for glucuronides of those compounds (Benet et al., 1993).
The study of the effects of isoform-specific inhibitors shown in Fig. 6 indicates that the glucuronidation of BVM by HLMs is not dependent on a single isozyme, even though recombinant UGT1A3 exhibited a distinctly higher rate (Fig. 3; Table 1). Thus, even very high concentrations of putative competitive inhibitors will not be likely to have a significant effect in vivo. This low potential for competitive drug-drug interactions is common for UGT substrates such as BVM that use multiple UGT isoforms and have fairly low or moderate binding affinities (Williams et al., 2004). The lack of an inhibitory effect of either bilirubin or etoposide, both UGT1A1 substrates, suggests that BVM may have little effect on bilirubin metabolism in vivo.

Inhibitory effects of typical probe substrates on BVM UGT activity in HLMs. BVM UGT activities at 25 μM BVM were determined as described under Materials and Methods. Bilirubin and etoposide (UGT1A1), imipramine (UGT1A3 and UGT1A4), trifluoperazine (UGT1A4), AZT, and diclofenac (UGT2B7) were used as inhibitors with the concentration ranges of 0 to 500 μM. Data (means of individual duplicate incubations) are expressed as peak area ratios relative to internal standard (DSD) in comparison with the controls without inhibitors. A, mono-BVMG (I); B, mono-BVMG (II).
Human UGTs generally exhibit distinct but overlapping substrate specificities and most substrates are glucuronidated by more than one isoform (Radominska-Pandya et al., 1999; King et al., 2000; Ritter, 2000; Tukey and Strassburg, 2000; Fisher et al., 2001). To our knowledge, the selectivity of UGT1A3 toward carboxylic acid-containing compounds, either aliphatic or aromatic carboxylic acids, has not been previously reported. UGT1A3 has been shown to catalyze the glucuronidation of some carboxylic acids, such as NSAIDs (e.g., naproxen and ibuprofen) and fibrates (e.g., ciprofibrate). However, it exhibited very low activity and no specific efficiency for these substrates (Green and Tephly, 1998; Sallustio et al., 2000; Sakaguchi et al., 2004). It has been reported that UGT1A3 is the specific isoform that catalyzes the glucuronidation of coumarin-3-carboxylic acid (Sakaguchi et al., 2004), but the formation rate, as measured by radioactive thin-layer chromatography, was very low (13–16 pmol/min/mg of protein). In addition, other UGT isoforms such as UGT2B4, -2B15, and -2B17 were not investigated in that study. The glucuronidation of hexafluoro-1α,23S,25-trihydroxyvitamin D3, an oxidative metabolite of hexafluoro-1α,25-dihydroxyvitamin D3, was recently shown to be catalyzed mainly by UGT1A3, and, for this substrate, UGT1A4, -2B4, and -2B7 were also involved in its glucuronidation at lower activities (Kasai et al., 2005). Hexafluoro-1α,23S,25-trihydroxyvitamin D3 has multiple functional hydroxyl groups in its chemical structure; thus, possible positional glucuronidation isomers could not be excluded. Because the distinctive acyl glucuronidation of BVM, specifically mono-BVMG (I), is most efficiently catalyzed by UGT1A3, BVM might be considered as a potential selective substrate or inhibitor for UGT1A3, as few specific substrates for UGT1A3 have been reported to date.
In summary, the glucuronidation of BVM, a novel maturation inhibitor of HIV, at its two carboxylic acids occurs in both human liver and intestinal tissues. Up to three human UGT isoforms may contribute to the glucuronidation, with UGT1A3 being the most efficient for the formation of mono-BVMG (I), assuming expression of isoforms is similar in recombinant UGTs, whereas only UGT2B7 was demonstrated to form mono-BVMG (II) in vitro. At present, because of the limited knowledge of the relative expression of UGT isoforms in human tissues, we cannot unambiguously determine whether any of these UGT isoforms may be dominant in vivo for BVM conjugation. The apparent participation of several UGT isoforms in BVM glucuronidation and the low to moderate affinities do, however, suggest that possible drug-drug interactions with specific UGTs would be less likely to substantially influence in vivo disposition of BVM in humans.
Footnotes
-
This research was supported by a contract from Panacos Pharmaceuticals (to P.C.S.) and, in part, by National Institutes of Health Grant AI33066 from National Institute of Allergy and Infectious Diseases (to K.H.L.).
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.106.012815.
-
ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; NSAID, nonsteroidal anti-inflammatory drug; BVM, bevirimat, 3-O-(3′,3′-dimethylsuccinyl)-betulinic acid; HIV, human immunodeficiency virus; LC, liquid chromatography; ESI, electrospray ionization; MS, mass spectrometry; BVMG, glucuronide of BVM; HLM, human liver microsome; HIM, human intestinal microsome; DSD, 3-O-(3′,3′-dimethylsuccinyl)-dihydrobetulinic acid; UDPGA, uridine 5′-diphosphoglucuronic acid; d-SL, d-saccharic acid 1,4-lactone; PMSF, phenylmethylsulfonyl fluoride; TFA, trifluoroacetic acid; AZT, zidovudine; TFP, trifluoperazine; 7-HFC, 7-hydroxy-4-trifluoromethylcoumarin; 7-HFCG, 4-trifluoromethyl-7-hydroxycoumarin glucuronide; RLM, rat liver microsome; MLM, mouse liver microsome; DLM, dog liver microsome; MMLM, marmoset liver microsome; RIM, rat intestinal microsome; ACN, acetonitrile; HPLC, high-performance liquid chromatography; SPE, solid-phase extraction; HAc, glacial acetic acid; TFPG, trifluoperazine glucuronide.
- Received September 8, 2006.
- Accepted December 5, 2006.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}