


Abstract
Precision-cut human liver slices obtained from 11 donors were cultured for 72 h in a defined medium (serum free Williams' medium E) supplemented with 0.1 μM insulin and 0.1 μM dexamethasone (DEX). Liver slices were treated with 50 μM concentrations of β -naphthoflavone (BNF), lansoprazole, rifampicin (RIF), DEX and methylclofenapate and 500 μM sodium phenobarbital (NaPB). The relative apoprotein levels of 12 cytochrome P450 (P450) enzymes were determined in liver slice microsomes using a panel of antipeptide antibodies. Treatment with BNF significantly induced mean levels of CYP1A2 apoprotein to 160% of levels in 72-h control (no test compound) human liver slice microsomes. NaPB significantly induced levels of CYP3A4 apoprotein to 255% of control and RIF significantly induced levels of CYP2C19 and CYP3A4 apoproteins to 265 and 330% of control, respectively. In addition, treatment with RIF increased levels of CYP2A6 apoprotein to 205% of control, and treatment with both NaPB and RIF increased levels of CYP2B6 apoprotein to 370 and 615% of control, respectively. However, these increases were not statistically significant, owing to a variable response between liver slice preparations from different subjects, this being apparent for all inducible P450s. In contrast, none of the compounds examined significantly increased levels of CYP2C8, CYP2C9, CYP2D6, CYP2E1, and CYP4A11 apoproteins. Levels of CYP1A1 apoprotein were not detected in any liver slice sample, either before or after treatment with the model inducers. Overall, these results demonstrate the utility of cultured human liver slices for assessing the effects of chemicals on P450 enzymes.
Hepatic cytochrome P450 (P4501) enzymes are known to have a major role in the metabolism of both xenobiotics (including therapeutic agents) and certain endogenous compounds (Nelson et al., 1996; Parkinson, 1996; Pelkonen et al., 1998). In the development of new therapeutic agents, it is important to ascertain whether the compound will be an inducer or an inhibitor of P450 enzymes, to minimize potential drug interactions (Lin and Lu, 1998; Pelkonen et al., 1998; Tucker et al., 2001). While P450 inhibition studies are normally performed with either human liver microsomes or cDNA-expressed P450 enzymes, cultured human hepatocytes have been extensively used for P450 induction studies (Maurel, 1996; Li et al., 1997; Tucker et al., 2001). P450 induction in cultured hepatocytes may be assessed by a variety of techniques including measurement of mRNA levels, apoprotein levels and monooxygenase activities. By employing such techniques, the induction of P450 enzymes in the CYP1A, CYP2A, CYP2B, CYP2C, CYP2E, and CYP3A subfamilies in cultured human hepatocytes has been reported by a number of laboratories (Curi-Pedrosa et al., 1994; Donato et al., 1995;Chang et al., 1997; Kostrubsky et al., 1999; Meunier et al., 2000;Rodrı́guez-Antona et al., 2000; Sahi et al., 2000;Gerbal-Chaloin et al., 2001).
A number of studies have demonstrated that precision-cut liver slices may be an alternative in vitro liver model system to isolated hepatocytes. Liver slices maintain tissue architecture, so that all cell types are present, and like hepatocytes have been extensively used for studies of xenobiotic metabolism and xenobiotic-induced toxicity (Bach et al., 1996; Lerche-Langrand and Toutain, 2000). In contrast to cultured human hepatocytes, relatively few investigations have examined the use of human liver slices for P450 enzyme induction studies. The majority of these studies have focused on the induction of CYP1A enzymes, as determined by effects on marker enzyme activities, apoprotein levels, and/or mRNA levels. For example, Aroclor 1254, β-naphthoflavone (BNF), 3,3′-diindolylmethane, omeprazole, and 2,3,7,8-tetrachlorodibenzo-p-dioxin have been reported to induce CYP1A enzymes in cultured human liver slices (Lake et al., 1996,1998; Olson et al., 1997; Drahushuk et al., 1998; Glöckner et al., 1999). In one study rifampicin (RIF) was reported to induce CYP3A4 (Lake et al., 1997), and in a recently published abstract, cyclophosphamide and phenobarbital were reported to induce CYP2B6 and CYP3A4 in cultured human liver slices (Martin et al., 2002).
The aim of this study was to examine the inducibility of the major hepatic P450 enzymes, namely members of the CYP1A, CYP2B, CYP2C, CYP2D, CYP2E, CYP3A, and CYP4A subfamilies, in cultured human liver slices. Unlike previous studies (Lake et al., 1996, 1997, 1998; Drahushuk et al., 1998; Glöckner et al., 1999), where relatively small numbers (up to six preparations) of human liver slices were evaluated, in the present study liver slice preparations from 11 donors were examined. The model inducers selected comprised BNF, dexamethasone (DEX), lansoprazole (LANS), RIF, and sodium phenobarbital (NaPB), which are all known to induce various P450 enzymes in human hepatocytes (Curi-Pedrosa et al., 1994; Donato et al., 1995; Maurel, 1996; Chang et al., 1997; Pelkonen et al., 1998; Meunier et al., 2000; Gerbal-Chaloin et al., 2001). Human liver slices were also treated with methylclofenapate (MCP), which is a potent rodent peroxisome proliferator and CYP4A enzyme inducer (Lake, 1995). In keeping with their known in vivo properties, MCP and other rodent peroxisome proliferators have been shown to produce peroxisome proliferation, induction of peroxisomal fatty acid oxidizing enzyme and levels of CYP4A apoproteins in cultured rat liver slices (Beamand et al., 1993;Lake et al., 1996).
Precision-cut liver slices were treated with the test compounds for 72 h, as this treatment period has been previously shown to be suitable to observe induction of P450 enzyme activities and apoprotein levels in cultured human liver slices (Lake et al., 1996, 1997, 1998). Levels of CYP1A1, CYP1A2, CYP2A6, CYP2B6, CYP2D6, CYP2E1, CYP3A4, CYP3A5, and CYP4A11 apoproteins were quantified by immunoblotting with a panel of fully characterized monospecific antipeptide antibodies (Edwards et al., 1998). In addition, levels of CYP2C enzymes were quantified with an antibody that binds to CYP2C8, CYP2C9, and CYP2C19, these P450 forms being readily separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Edwards et al., 1998). Finally, limited studies were also performed with some enzymatic markers, namely 7-ethoxyresorufin O-deethylase, 7-ethoxy-4-trifluormethylcoumarin O-deethylase, and testosterone 6β-hydroxylase.
Materials and Methods
Chemicals.
The sources of the tissue culture materials were as described previously (Beamand et al., 1993). BNF, DEX, LANS, NaPB, RIF, α -naphthoflavone, 7-ethoxyresorufin, testosterone, resorufin, 6β-hydroxytestosterone, enzyme cofactors, prestained molecular weight markers, and protein G coupled to horseradish peroxidase were purchased from Sigma-Aldrich Co. Ltd. (Poole, Dorset, UK). MCP was purchased from Lancaster Synthesis (Morecambe, Lancs, UK) and 7-ethoxy-4-trifluoromethylcoumarin and 7-hydroxy-4-trifluoromethylcoumarin from Enzyme System Products (Livermore, CA). [4-14C]Testosterone (specific activity 56 mCi/mmol) was obtained from Amersham Pharmacia Biotech UK Ltd. (Little Chalfont, Buckinghamshire, UK) and SDS–polyacrylamide gel electrophoresis reagents from National Diagnostics (Aylesbury, Bucks, UK).
Preparation of Liver Slices.
Samples of human liver were collected and transported to BIBRA on ice (BIBRA International Ltd., Carshalton, Surrey, UK. The donors of the 11 human liver samples, designated subjects A to K, were female subjects aged 31, 38, 55, and 11 years, male subjects aged 13, 50, 10, 24, 25, and 66 years, and a female subject aged 17 years, respectively. The Research Ethics Committee of King's College Hospital and the donor's representatives had granted approval for the use of this tissue, which was surplus to clinical requirements. Tissue cylinders from liver samples were prepared using a 10-mm diameter motor-driven tissue-coring tool. From the cylinders, tissue slices (200–300 μm) were prepared in cold oxygenated (95% O2/5% CO2) Earle's balanced salt solution containing 25 mM d-glucose, 50 μg/ml gentamicin, and 2.5 μg/ml fungizone using a Krumdieck tissue slicer (Alabama Research and Development Corp., Munford, AL).
Culture of Liver Slices.
Liver slices were floated onto Vitron Inc. (Tucson, AZ) type C titanium roller inserts (two slices per insert) and cultured at 2 rpm in glass vials containing 1.7 ml of culture medium employing a Vitron dynamic organ culture incubator, which was operated in accordance with the manufacturers' instructions. The culture medium consisted of serum free Williams' medium E containing 2 mM l-glutamine, 0.1 μM insulin, 0.1 μM DEX, 50 μg/ml gentamicin, and 2.5 μg/ml fungizone. Liver slice cultures were maintained at 37°C in an atmosphere of 95% O2/5% CO2. After 1 h, treatment was commenced by replacing the culture medium with fresh medium containing the required concentrations of the test compounds. BNF, DEX, LANS, MCP, and RIF were dissolved in dimethyl sulfoxide (DMSO) prior to addition to the culture medium, whereas NaPB was dissolved directly in the culture medium. The final DMSO concentration was 0.4% (v/v) in all vials including the control cultures. Subsequently, the medium was replaced every 24 h and the slices redosed with fresh medium containing the test compounds. Following each medium change and returning the vials to the incubator, the gas flow was increased for 10 s to restore the high oxygen atmosphere.
Biochemical Investigations with Liver Slices.
Freshly cut and 72-h cultured liver slices were washed in ice-cold 0.154 M KCl containing 50 mM Tris-HCl, pH 7.4, homogenized in this medium by sonication (Beamand et al., 1993), and stored at −80°C. Liver slice homogenates (prepared from 16–20 slices) were centrifuged at 10,000g average for 15 min to obtain the postmitochondrial supernatant fraction and subsequently at 158,000g average for 40 min to separate the microsomal fraction from the cytosol. Microsomal fractions were resuspended in 0.154 M KCl containing 50 mM Tris-HCl, pH 7.4, and 2 mM EDTA and aliquots stored at −80°C. Protein was determined as described previously (Renwick et al., 2000). In some experiments liver slice unwashed microsomal fractions were assayed for activities of 7-ethoxyresorufin O-deethylase and testosterone 6β-hydroxylase as described previously (Renwick et al., 2000). The activity of 7-ethoxy-4-trifluoromethylcoumarin O-deethylase was determined directly at 37°C in spectrofluorimeter cuvettes containing 5 μM substrate (added in 5 μl DMSO), 5 μM α-naphthoflavone (added in 5 μl DMSO), 1 mM NADP+, 7.5 mM dl-isocitric acid, 1 unit/ml isocitric dehydrogenase, 5 mM MgSO4, 0.15 to 0.20 mg microsomal protein and 100 mM phosphate buffer, pH 7.4, in a final volume of 2 ml. 7-Hydroxy-4-trifluoromethylcoumarin formation was monitored at wavelengths of 410 nm excitation and 510 nm emission. α-Naphthoflavone was added to inhibit the contribution of CYP1A2 to this reaction (Code et al., 1997).
Immunoblotting.
Immunoblotting was performed using 10 to 100 μg of liver slice unwashed microsomal protein, as appropriate, as described previously (Renwick et al., 2000). The levels of CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2D6, CYP2E1, CYP3A4, CYP3A5, and CYP4A11 were determined as amounts relative to those present in a pooled microsomal preparation of six normal human livers. The relative levels of CYP2C19 were assessed by using a single sample of human liver microsomes that contained a readily detectable amount of CYP2C19. For CYP1A1, a sample of microsomes from baculovirus-infected insect cells containing cDNA-expressed CYP1A1 (BD Gentest, Woburn, MA; obtained from Cambridge Bioscience, Cambridge, UK) was used as reference. Immunoreactive band intensity was quantified using a Kodak Image Station and Kodak Digital Science ID Image analysis software (PerkinElmer Life Sciences, Hounslow, UK).
Statistical Analysis.
The data were analyzed using Student's paired t test.
Results
Effect of Model Inducers on Levels of P450 Apoproteins.
Immunoblotting of freshly cut and 72-h cultured human liver slice microsomes was performed to assess the relative levels of CYP1A1, CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5, and CYP4A11 apoproteins. Because of the small amounts of material available, it was not possible to examine the effects of all of the inducers studied on liver slices from every donor. However, the effects of each of the model inducers were determined on levels of all the P450 apoproteins evaluated in this study.
As reported previously (Renwick et al., 2000), levels of P450 apoproteins declined in human liver slices cultured for 72 h in control medium. For all the experiments conducted, mean levels of CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5, and CYP4A11 apoproteins in 72-h cultured liver slices were around 20 to 80% of levels in freshly cut human liver slices (data not shown).
In samples from some donors, one or more P450 apoproteins were not detectable in freshly cut human liver slice microsomes. Where P450 apoproteins were absent from freshly cut liver slices, they remained undetectable in 72-h cultured liver slices treated with any of the model inducers studied. Consequently, such samples were excluded from the statistical analysis of the inducibility of these P450s.
Human liver slices were cultured for 72 h with 50 μM BNF, 50 μM DEX, 50 μM LANS, 50 μM MCP, 500 μM NaPB, and 50 μM RIF. The concentrations of BNF, DEX, LANS, NaPB, and RIF were selected from previous in vitro studies in either human hepatocytes (Curi-Pedrosa et al., 1994; Donato et al., 1995; Meunier et al., 2000; Sahi et al., 2000) or liver slices (Lake et al., 1996, 1997). For MCP, a rodent peroxisome proliferator and CYP4A inducer, the concentration selected was based on that used in previous studies with cultured rat and human liver slices (Beamand et al., 1993; Lake et al., 1996). Because of limited tissue availability, no studies were undertaken to determine the optimal concentrations of the compounds examined for P450 induction studies in cultured human liver slices.
Levels of immunoreactive CYP1A1 were not detected in any experiment using microsomes from freshly cut slices, 72-h control (no test compound treated) slices or from slices cultured for 72 h with any of the model inducers studied.
Treatment with BNF significantly increased mean CYP1A2 apoprotein levels to 160%, of those in 72-h cultured control liver slice microsomes (Fig. 1A). In contrast, treatment with LANS, NaPB, RIF, DEX, and MCP had no significant effect on CYP1A2 apoprotein levels in cultured human liver slices.

Effect of treatment of human liver slices with BNF, LANS, NaPB, RIF, DEX, and MCP for 72 h on levels of CYP1A2 (A), CYP2A6 (B), and CYP2B6 (C) apoproteins.
Microsomes were prepared from liver slices cultured for 72 h in either control medium (DMSO only treated) or medium containing the model inducers (concentrations 500 μM for NaPB and 50 μM for the other compounds) and immunoblotting performed. Results are presented as mean ± S.E. with the number of subjects studied shown in parentheses for relative P450 apoprotein levels (see Materials and Methods) in 72-h cultured liver slices. Values significantly different from control are ∗, p < 0.05.
Although treatment of human liver slices with RIF increased mean levels of CYP2A6 apoprotein to 205% of those in 72-h cultured control liver slice microsomes (Fig. 1B) and treatment with both NaPB and RIF increased mean levels of CYP2B6 apoprotein (Fig. 1C) to 370 and 615%, respectively, none of these increases reached statistical significance. However, it is notable that there was a considerable variability in the response to the inducers (see below). None of the other model inducers examined had any significant effect on levels of CYP2A6 and CYP2B6 apoproteins in 72-h cultured human liver slices.
None of the model inducers examined had any marked effect on CYP2C8 apoprotein levels (Fig. 2A). Whereas none of the model inducers significantly increased levels of CYP2C9 apoprotein, treatment with MCP resulted in a significant reduction of mean CYP2C9 apoprotein levels to 45% of 72-cultured control liver slice values (Fig. 2B). In contrast, RIF significantly increased mean levels of CYP2C19 apoprotein to 265% of 72-h control liver slice values (Fig. 2C).

Effect of treatment of human liver slices with BNF, LANS, NaPB, RIF, DEX, and MCP for 72 h on levels of CYP2C8 (A), CYP2C9 (B), and CYP2C19 (C) apoproteins.
Microsomes were prepared from liver slices cultured for 72 h in either control medium (DMSO only treated) or medium containing the model inducers (concentrations 500 μM for NaPB and 50 μM for the other compounds) and immunoblotting performed. Results are presented as either mean ± S.E. or mean with the number of subjects studied shown in parentheses for relative P450 apoprotein levels (seeMaterials and Methods) in 72-h cultured liver slices. Values significantly different from control are ∗,p < 0.05.
Treatment of human liver slices for 72 h with BNF, NaPB, RIF, DEX, and MCP did not significantly increase mean levels of CYP2D6 (Fig.3A) and CYP2E1 (Fig. 3B) apoproteins. Whereas treatment with LANS did not significantly increase levels of CYP2D6 apoprotein (Fig. 3A), mean levels of CYP2E1 apoprotein were significantly reduced to 65% of 72-h control liver slice values (Fig.3B).

Effect of treatment of human liver slices with BNF, LANS, NaPB, RIF, DEX, and MCP for 72 h on levels of CYP2D6 (A) and CYP2E1 (B) apoproteins.
Microsomes were prepared from liver slices cultured for 72 h in either control medium (DMSO only treated) or medium containing the model inducers (concentrations 500 μM for NaPB and 50 μM for the other compounds) and immunoblotting performed. Results are presented as mean ± S.E. with the number of subjects studied shown in parentheses for relative P450 apoprotein levels (see Materials and Methods) in 72-h cultured liver slices. Values significantly different from control are ∗, p < 0.05.
Treatment of human liver slices for 72 h with BNF, LANS, DEX, and MCP had no significant effect on mean levels of CYP3A4 apoprotein (Fig.4A). In contrast, treatment with NaPB and RIF both significantly increased mean levels of CYP3A4 apoprotein to 255 and 330% of 72-h control liver slice values, respectively. However, neither NaPB nor RIF had any significant effect on levels of CYP3A5 apoprotein in 72-h cultured human liver slices (Fig. 4B). While treatment with LANS, DEX, and MCP had no significant effect on levels of CYP3A5 apoprotein, treatment with BNF produced a small, but statistically significant, increase in CYP3A5 apoprotein levels.

Effect of treatment of human liver slices with BNF, LANS, NaPB, RIF, DEX, and MCP for 72 h on levels of CYP3A4 (A), CYP3A5 (B), and CYP4A11 (C) apoproteins.
Microsomes were prepared from liver slices cultured for 72 h in either control medium (DMSO only treated) or medium containing the model inducers (concentrations 500 μM for NaPB and 50 μM for the other compounds) and immunoblotting performed. Results are presented as mean ± S.E. with the number of subjects studied shown in parentheses for relative P450 apoprotein levels (see Materials and Methods) in 72-h cultured liver slices. Values significantly different from control are ∗, p < 0.05; ∗∗, p < 0.01.
The treatment of human liver slices for 72 h with BNF, LANS, NaPB, RIF, and DEX had no significant effect on mean levels of CYP4A11 apoprotein (Fig. 4C). In contrast, treatment with the rodent peroxisome proliferator MCP resulted in a significant decrease in mean CYP4A11 apoprotein levels to 60% of 72-h control liver slice values.
Variability in Induction of P450 Apoproteins in Human Liver Slices.
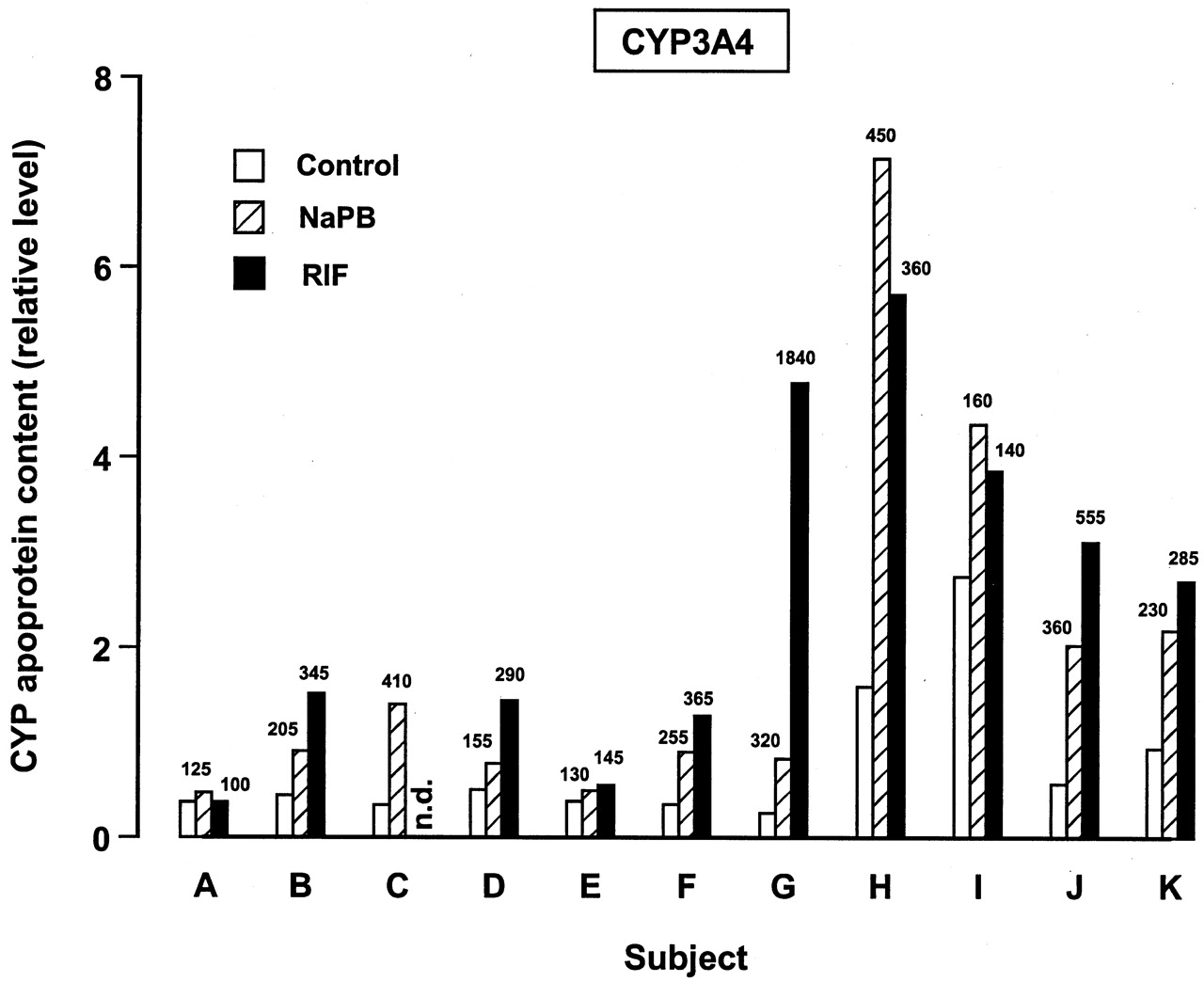
For all the inducible P450 enzymes, variability was observed among individual donors in the magnitude of the response to the model inducers. This variability is illustrated in Figs.5 and 6 for the effect of NaPB and RIF on levels of CYP3A4 and CYP2B6 apoproteins. Figure 5 shows the effect of NaPB and RIF on levels of CYP3A4 apoprotein in 72-h cultured liver slices from each individual subject. The effect of NaPB was studied in liver slices from all eleven subjects, whereas the effect of RIF was evaluated in liver slices from all but one subject (subject C). Whereas little response to the two model inducers was observed in liver slice preparations from some subjects (e.g., subjects A and E), a marked response to both NaPB and RIF was observed with liver slices from other subjects (e.g., subjects G, H, and J). Overall, CYP3A4 apoprotein levels following treatment with NaPB and RIF in these studies ranged from 125 to 450 and from 100 to 1840% of control, respectively (Fig. 5). The effect of NaPB and RIF was evaluated with five subjects where the liver slice preparations contained detectable levels of CYP2B6 apoprotein (Fig. 6). Compared with levels of CYP2B6 apoprotein in 72-h cultured control liver slice microsomes, both model inducers produced ≥2-fold increases in CYP2B apoprotein levels. For these human liver slice preparations, following treatment with NaPB and RIF CYP2B6 apoprotein levels ranged from 200 to 1740 and from 310 to 1395% of control, respectively (Fig. 6).
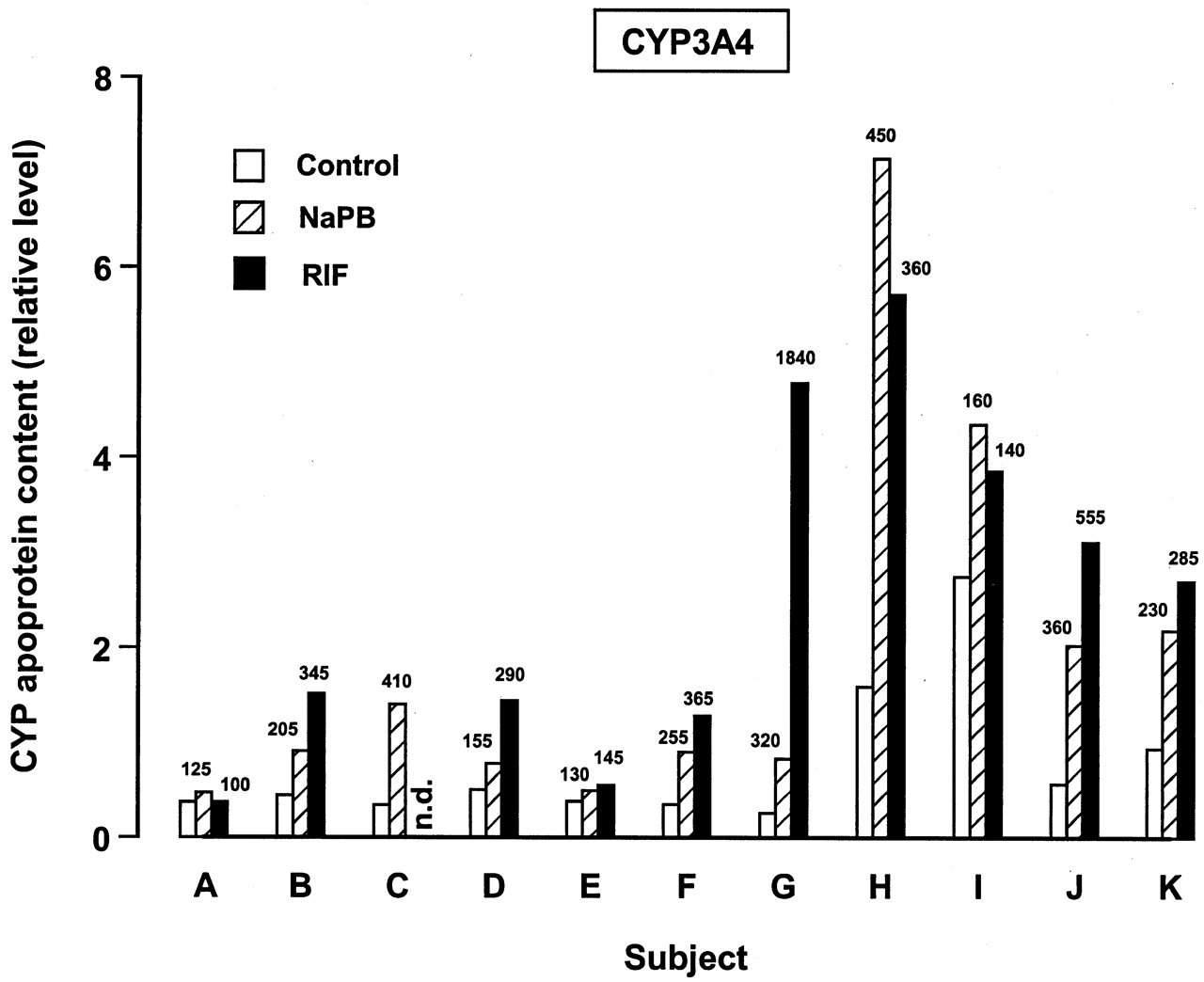
Variability in the effect of treatment of human liver slices with NaPB and RIF for 72 h on levels of CYP3A4 apoprotein.
Microsomes were prepared from liver slices cultured for 72 h in control medium (DMSO only treated) and medium containing either 500 μM NaPB or 50 μM RIF and immunoblotting performed. Results are presented for individual experiments for relative P450 apoprotein levels (see Materials and Methods), with percentage of levels in 72-h cultured control liver slices following treatment with NaPB and RIF also being shown (numbers above bars). The effect of RIF was not evaluated (n.d. = not determined) with liver slices from subject C.

Variability in the effect of treatment of human liver slices with NaPB and RIF for 72 h on levels of CYP2B6 apoprotein.
Microsomes were prepared from liver slices cultured for 72 h in control medium (DMSO only treated) and medium containing either 500 μM NaPB or 50 μM RIF and immunoblotting performed. Results are presented for individual experiments for relative P450 apoprotein levels (see Materials and Methods), with percentage of levels in 72-h cultured control liver slices following treatment with NaPB and RIF also being shown (numbers above bars).
Effect of Model Inducers on Levels of CYP1A2, CYP2B6, and CYP3A4 Enzyme Activities and Apoproteins.
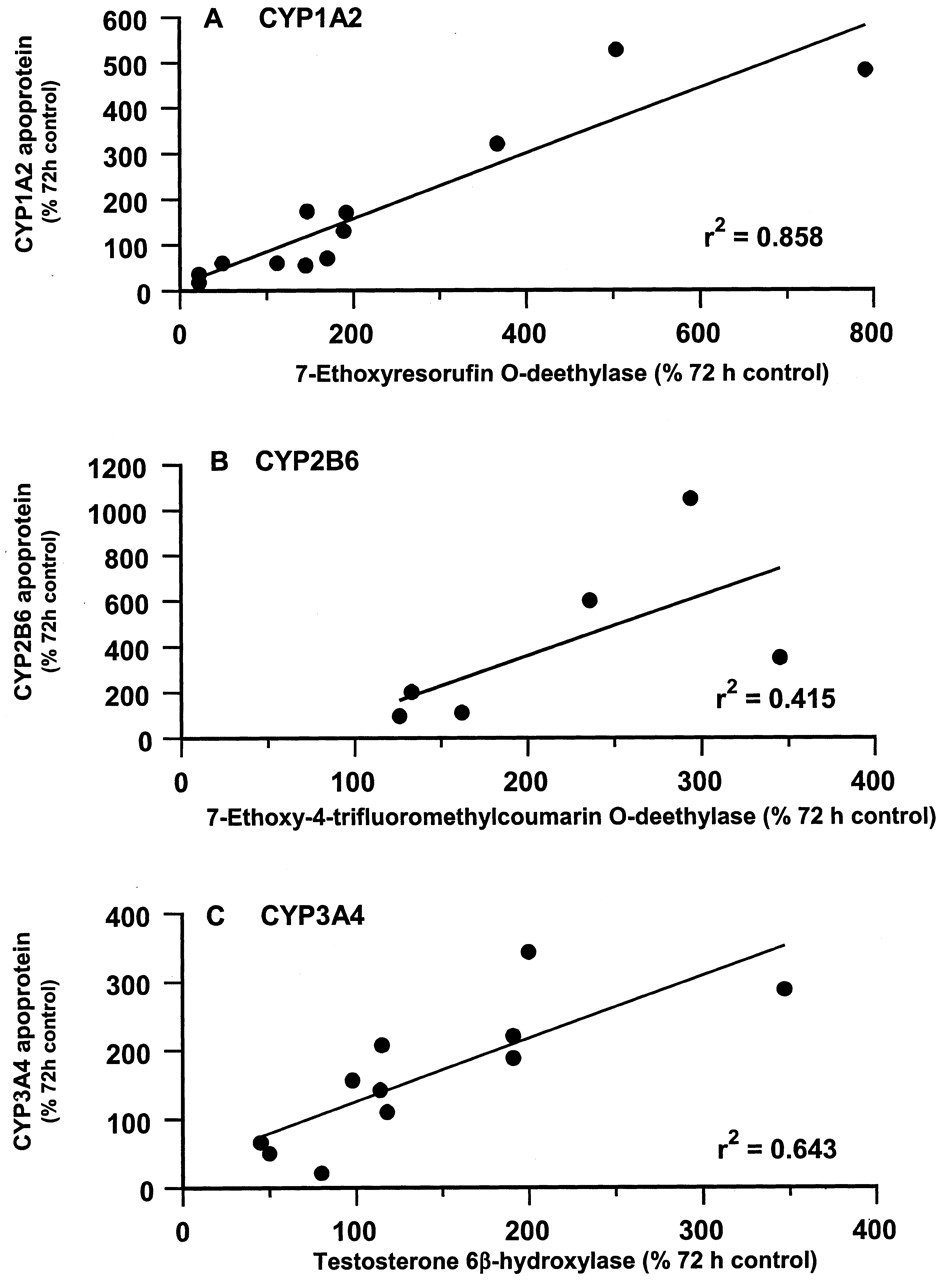
In some experiments there was sufficient material available from a donor sample to determine enzymatic markers for CYP1A2, CYP2B6, and CYP3A4 in microsomes from liver slices cultured for 72 h with BNF, LANS, NaPB, RIF, DEX, and MCP. The activities of 7-ethoxyresorufinO-deethylase, 7-ethoxy-4-trifluoromethylcoumarinO-deethylase and testosterone 6β-hydroxylase (all expressed as percentage of 72-h control human liver slice microsome levels) were then compared with levels of their respective P450 apoproteins (normalized to their respective 72-h control values). Good correlations were observed between 7-ethoxyresorufin O− deethylase activity and CYP1A2 apoprotein levels (r2 = 0.858; Fig.7A) and between testosterone 6β-hydroxylase activity and CYP3A4 apoprotein levels (r2 = 0.643; Fig. 7C).

Correlation between levels of CYP1A2 (A), CYP2B6 (B), and CYP3A4 (C) apoproteins and associated enzyme activities in human liver slices treated with BNF, LANS, NaPB, RIF, DEX, and MCP for 72 h.
Microsomes were prepared from liver slices cultured for 72 h in either control medium (DMSO only treated) or medium containing the model inducers (concentrations 500 μM for NaPB and 50 μM for the other compounds) and immunoblotting and enzyme activity determinations performed as described under Materials and Methods. Results are presented for experiments with two subjects for CYP1A2 (subjects D and F) and CYP3A4 (subjects B and D) and one subject for CYP2B6 (subject D). P450 apoprotein levels and enzyme activities are expressed as a percentage of levels in 72-h cultured control liver slices.
In the one experiment performed, a lower correlation (r2 = 0.415; Fig. 6B) was observed between 7-ethoxy-4-trifluoromethylcoumarin O-deethylase activity and levels of CYP2B6 apoprotein. The lower correlation between 7-ethoxy-4-trifluoromethylcoumarin O-deethylase activity and CYP2B6 apoprotein levels may be attributable to the observation that even in the presence of α-naphthoflavone to inhibit CYP1A2 activity, this substrate is not specific for CYP2B6, being also metabolized by other P450 enzymes in human liver (Bogaards et al., 1996; Code et al., 1997).
Discussion
The aim of the current study was to examine the usefulness of cultured precision-cut human liver slices as an in vitro liver model system for P450 induction studies. By employing a characterized panel of highly specific antipeptide antibodies (Edwards et al., 1998), the effect of a range of model inducers on the levels of twelve P450 apoproteins in 72-h culture human liver slices was determined. At the concentrations examined, BNF, NaPB, and RIF all significantly increased levels of one or more P450 apoproteins in cultured human liver slices. Whereas the majority of this work was conducted by measuring apoprotein levels, some limited studies were also performed with enzymatic markers. Good correlations were obtained between levels of CYP1A2 and CYP3A4 apoproteins and their respective enzymatic markers, suggesting that the increased levels of apoproteins observed after treatment with the model inducers were catalytically active.
The treatment of human liver slices with BNF resulted in an increase in CYP1A2 apoprotein levels, presumably via an aryl-hydrocarbon receptor-dependent mechanism. This is consistent with the results of studies on the effects of BNF on CYP1A2 in cultured human hepatocytes by some workers (Curi-Pedrosa et al., 1994; Meunier et al., 2000; Sahi et al., 2000) but not by others (Liu et al., 2001). In the present study, CYP1A1 apoprotein levels remained below the limit of detection, even after treatment with BNF and LANS. This contrasts with results of some previous studies with human hepatocytes (Curi-Pedrosa et al., 1994; Allen et al., 2001; Liu et al., 2001) and liver slices (Drahushuk et al., 1998). However, the differential induction of CYP1A apoproteins in the present study is entirely consistent with the response of such inducers in vivo (Sesardic et al., 1990). The absence of detectable levels of CYP1A1 apoprotein is in agreement with other studies using this panel of antipeptide antibodies performed both in cultured human liver slices (Renwick et al., 2000) and freshly isolated liver microsomes (Edwards et al., 1998). There is evidence that hepatic CYP1A1 in humans is subject to negative regulation (Piechocki and Hines, 1998), whereas CYP1A2 is constitutively expressed and inducible in adult liver (Pelkonen et al., 1998).
Previous studies have demonstrated that NaPB and RIF can induce a number of P450 enzymes in human hepatocytes (Curi-Pedrosa et al., 1994;Chang et al., 1997; Meunier et al., 2000; Rodrı́guez-Antona et al., 2000; Sahi et al., 2000; Gerbal-Chaloin et al., 2001). In the present study, NaPB significantly increased levels of CYP3A4 apoprotein, and RIF significantly increased levels of CYP2C19 and CYP3A4 apoproteins. Treatment with RIF appeared to increase levels of CYP2A6 apoprotein in some subjects and treatment with both NaPB and RIF increased levels of CYP2B6 apoprotein in all subjects where constitutive levels of CYP2B6 were detectable. However, these increases were not statistically significant, although the type II error was quite large due to the limited number of human liver slice preparations that expressed these P450 apoproteins at detectable levels and the variability in response between individual subjects. The effects of NaPB and RIF on CYP2B6 and CYP3A4 were qualitatively similar to those observed in a previous study where mRNA levels were quantified in cultured human hepatocytes (Goodwin et al., 2001). Additional studies are required to fully evaluate the effects of NaPB and RIF on levels of CYP2A6 and CYP2B6 apoproteins in cultured human liver slices.
In terms of the overall mean values for all the P450s examined, both NaPB and RIF produced the largest increases in levels of CYP2B6 apoprotein. These results with cultured liver slices support previous studies with hepatocytes (Chang et al., 1997; Rodrı́guez-Antona et al., 2000; Sahi et al., 2000; Gerbal-Chaloin et al., 2001), that this P450 enzyme is markedly inducible in human liver. Although both NaPB and RIF significantly increased levels of CYP3A4 apoprotein, neither compound produced a significant increase in levels of CYP3A5 apoprotein, suggesting that this P450 enzyme is less responsive than CYP3A4 to model inducers. Similar observations have been reported in cultured human hepatocytes (Rae et al., 2001).
The effects of DEX on P450 expression in human hepatocytes are complex, due in part to the physiological role of glucocorticoids in maintaining expression of the pregnane X receptor and the constitutive androstane receptor (Pascussi et al., 2001). In the present studies, the fully defined culture medium was supplemented with 0.1 μM insulin and 0.1 μM DEX. At these levels it has been reported that DEX can enhance the induction of CYP2B6, CYP2C8, CYP2C9, and CYP3A4 mRNA levels in human hepatocytes by NaPB and RIF (Pascussi et al., 2001). At the 50 μM concentration examined in the present study, DEX had no significant effect on levels of P450 apoproteins in cultured human liver slices. However, in human hepatocytes, DEX has been shown to induce several P450 enzymes including CYP2A6, CYP2B6, CYP2C8, CYP2C9, and CYP3A4 (Donato et al., 1995; Chang et al., 1997; Meunier et al., 2000; Sahi et al., 2000; Gerbal-Chaloin et al., 2001). In these studies, the effects of DEX were examined over a wide range of concentrations from submicromolar up to 500 μM. Additional studies are required to evaluate the effects of a wide range of concentrations of DEX, including omitting DEX from the standard medium, on P450 enzymes in cultured human liver slices.
In recent years much progress has been made in elucidating the various pathways and receptors involved in the regulation of P450 enzymes in mammalian liver (Waxman, 1999; Honkakoski and Negishi, 2000;Gerbal-Chaloin et al., 2001). From the results obtained in this study, as with cultured hepatocytes, the various pathways for induction of CYP1A, CYP2A, CYP2B, CYP2C, and CYP3A subfamily P450 enzymes are retained in cultured human liver slices. In contrast to these P450 enzymes, none of the model inducers examined increased levels of CYP2D6, CYP2E1, and CYP4A11 apoproteins in 72-h cultured human liver slices. For CYP2D6, previous studies have suggested that this P450 enzyme is refractory to induction by known inducers of other P450 subfamilies (Parkinson, 1996; Rodrı́guez-Antona et al., 2000). Treatment with the rodent peroxisome proliferator MCP resulted in a reduction in the levels of liver slice CYP4A11 apoprotein. MCP had a similar effect on CYP2C9 apoprotein but had little effect on the other P450s examined. The effects of MCP thus appear to be selective and not due to a reduction in the functional viability of the human liver slice preparations. These results provide further evidence for species differences in the hepatic effects of rodent peroxisome proliferators (Lake, 1995).
For all of the inducible P450 enzymes examined in this study, marked variability in both the expression levels and the magnitude of induction was observed between liver slices from different donors. This variability is illustrated in Figs. 5 and 6 for the effect of NaPB and RIF on levels of CYP3A4 and CYP2B6 apoproteins. Despite this variability, both compounds increased CYP3A4 apoprotein levels in the majority of the subjects examined, with CYP2B6 apoprotein being increased in all samples where it could be detected. Variability in response to model inducers has also been observed in previous studies with cultured human hepatocytes (Curi-Pedrosa et al., 1994; Maurel, 1996; Meunier et al., 2000; Gerbal-Chaloin et al., 2001) and liver slices (Lake et al., 1997, 1998). Recently, Martin et al. (2002)suggested that the variability between subjects in CYP2B6 and CYP3A4 induction was less in human liver slices than in hepatocytes.
Such variability in response between subjects may be due to a number of factors. Due to genetic and environmental factors, the expression of CYP3A (Koch et al., 2002) and other P450 genes can vary considerably between subjects. In addition, for some of the P450s studied, polymorphisms are known to exist, and these will account for some of the nondetectable levels. Possibly, polymorphisms may also result in a reduced stability of P450 apoproteins. Another important factor is tissue quality. The extent to which this affects the induction response is not known, but it may be worthwhile including appropriate viability markers in liver slice P450 induction studies. For example, some workers have used a slice potassium content of 40 μmol/g or greater to assess the suitability of human liver slice preparations for CYP1A induction studies (Olson et al., 1997; Drahushuk et al., 1998).
Overall, the available data suggests that it is desirable to examine a number of human hepatocyte or liver slice preparations to assess the effects of a new chemical entity on P450 enzymes. It is important to assess baseline specific apoprotein content, to avoid poor metabolizesr (“null”) samples. Moreover, as recommended for P450 induction studies with human hepatocytes (Li et al., 1997; Tucker et al., 2001), suitable positive controls (e.g., BNF, NaPB, RIF) need to be included in P450 induction studies employing human liver slices, and replicate experiments should be performed. When assessing the effects of new compounds it may also be worthwhile to evaluate potential toxicity to the liver slices, by including appropriate viability markers in the experimental design.
In conclusion, these results demonstrate the utility of cultured human liver slices for assessing the effects of chemicals on P450 enzymes.
Footnotes
-
We gratefully acknowledge the provision of financial support by a grant from the Commission of the European Communities (Project “Eurocyp”, BIOMED Contract No. BMH4-CT96-0254) and a Realizing Our Potential Award from the Medical Research Council.
- Abbreviations used are::
- P450
- cytochrome P450
- BNF
- β-naphthoflavone
- RIF
- rifampicin
- DEX
- dexamethasone
- LANS
- lansoprazole
- MCP
- methylclofenapate
- NaPB
- sodium phenobarbital
- DMSO
- dimethyl sulfoxide
- Received August 26, 2002.
- Accepted November 26, 2002.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}