


Abstract
UDP-glucuronosyltransferase (UGT) 2B17 is one of the most important conjugating enzymes in androgen metabolism and shares more than 95% homology with UGT2B15. Although UGT2B15 has been fully characterized for its ability to conjugate drugs, environmental pollutants, and dietary components, UGT2B17 received less attention for its capacity to glucuronidate xenobiotics. In the present study, more than 55 exogenous compounds belonging to several categories of compounds were analyzed as potential substrates for UGT2B17. Glucuronidation activity was observed with several coumarins, anthraquinones, and flavonoids. The higher glucuronidation activity was measured with alizarin (125 pmol · min−1 · mg protein−1), whereas UGT2B17 conjugated eugenol, scopoletin, and galangin with glucuronidation rates of 102.5, 102, and 58 pmol · min−1 · mg protein−1, respectively. The characterization of UGT2B17 as a xenobiotics-conjugating enzyme demonstrates that its role is not limited to androgen metabolism and that its specificity for exogenous substrates is different from other UGT2B isoforms. Taken together, these data suggest a role of UGT2B17 for the hepatic detoxification.
Glucuronidation is one of the major metabolic pathways implicated in the elimination of exogenous compounds from organisms (Evans and Relling, 1999). This phase II metabolic reaction is catalyzed by UDP-glucuronosyltransferase (UGT2) enzymes, which transfer the glucuronosyl group from the UDP-glucuronic acid (UDPGA) to several lipophilic compounds such as bile acids, bilirubin, steroid hormones, environmental pollutants, drugs, and dietary components. The conjugated derivatives are more polar and thus more easily excreted through the bile or urine. Based on sequence homology, UGT proteins have been categorized into two families, UGT1 and UGT2 (Mackenzie et al., 1997). In humans, members of the UGT1A family are encoded by a complex gene localized on chromosome 2q37 (Owens and Ritter, 1995; Mackenzie et al., 1997). These enzymes glucuronidate bilirubin, amines, several drugs, and steroid hormones, especially estrogens and catecholestrogens (Senafi et al., 1994; King et al., 1996; Green et al., 1998; Green and Tephly, 1998; Albert et al., 1999). Members of the UGT2 family are encoded by distinct genes composed of six exons and located on chromosome 4q13-4q21.1 (Turgeon et al., 2000). Isoforms of the UGT2 family are further divided into two subfamilies, UGT2A enzymes metabolize molecules in the olfactive epithelium (Jedlitschky et al., 1999), whereas UGT2B proteins are expressed in large number of tissues, namely, liver, kidney, skin, brain, breast, uterine, and prostate (Mackenzie et al., 1997).
To date, seven UGT2B enzymes have been characterized: UGT2B4 (Fournel Gigleux et al., 1989; Lévesque et al., 1999), UGT2B7 (Ritter et al., 1990; Coffman et al., 1997), UGT2B10 (Jin et al., 1993), UGT2B11 (Beaulieu et al., 1998), UGT2B15 (Chen et al., 1993; Lévesque et al., 1997), UGT2B17 (Beaulieu et al., 1996), and UGT2B28 (Lévesque et al., 2001). Although substrates for UGT2B10 and UGT2B11 have not been identified, all other UGT2B isoforms conjugate various endogenous substances. UGT2B4 conjugates biliary acids, whereas UGT2B7, UGT2B15, UGT2B17, and UGT2B28 recognize dihydrotestosterone and its metabolites androstane-3α,17β-diol (3α-diol) and androsterone. Interestingly, it was suggested that these UGT2B enzymes play an important role in steroid target tissues to inactivate androgens (Turgeon et al., 2001). The classical role of these UGT2B isoforms in detoxification is also supported by their expression in the gastrointestinal tract, liver, and kidney (Coffman et al., 1997, 1998;Barbier et al., 2000; King et al., 2000; Turgeon et al., 2001). Indeed, several studies have previously demonstrated that UGT2B7 and UGT2B15 conjugate a large series of xenobiotics compounds (Green et al., 1994;Coffman et al., 1998). However, UGT2B17, an enzyme that shares more than 95% homology with UGT2B15, received less attention for its capacity to conjugate xenobiotics. In fact, this enzyme has been studied exclusively for its role in androgen inactivation (Beaulieu et al., 1996). Interestingly, UGT2B15 and UGT2B17 proteins are 95% identical in primary structure; however, UGT2B17 is capable of conjugating C19-steroid molecules at both the 3α- and 17β-OH positions, whereas UGT2B15 is only active at the 17β-OH position (Beaulieu et al., 1996). However, mutation of the serine residue to a tyrosine at position 121 of UGT2B17, as found in UGT2B15, abolished the ability of UGT2B17 to conjugate androgens at the 3α-position, but still retained activity for conjugation at the 17β position (Beaulieu et al., 1996; Lévesque et al., 1997; Dubois et al., 1999). These data indicate that minor changes in the primary structure of UGT2B enzymes have major influence in the substrate specificity.
To investigate substrate specificity of UGT2B17 toward exogenous compounds, coumarins, anthraquinones, flavonoids, and other classes of molecules were screened with microsomal proteins from human embryonic kidney 293 (HK293) cells stably transfected with the UGT2B17 cDNA. Our data demonstrate that UGT2B17 conjugates several members of almost all classes of molecules tested. As for steroids, it is demonstrated that UGT2B17 possesses distinct substrate specificity for the conjugation of exogenous compounds from other hepatic UGT enzymes, indicating that it plays an important role in xenobiotics inactivation and metabolism.
Materials and Methods
Materials.
UDP-glucuronic acid and all aglycon substrates were obtained from Sigma-Aldrich (St. Louis, MO) and ICN Pharmaceuticals (Montreal, QC, Canada). [14C]UDP-glucuronic acid (285 mCi/mmol) was obtained from PerkinElmer Life Sciences (Boston, MA). Geneticin (G418) and lipofectin were obtained from Invitrogen (Burlington, ON, Canada). Protein assay reagents were obtained from Bio-Rad (Hercules, CA). Restriction enzymes and other molecular biology reagents were from Pharmacia LKB Biotechnology Inc. (Milwaukee, WI), Invitrogen, Stratagene (San Diego, CA), and Roche Diagnostics (Indianapolis, IN). HK293 cells were obtained from the American Type Culture Collection (Manassas, VA).
Stable Expression of the UGT2B17 Enzyme.
HK293 cells were grown in Dulbecco's modified Eagle's medium containing 4.5 g/l glucose, 10 mM HEPES, 100 μg/ml sodium pyruvate, 100 units of penicillin/ml, 100 μg/ml streptomycin, and 10% (v/v) fetal bovine serum in a humidified incubator with an atmosphere of 5% CO2 and 37°C. The HK293-UGT2B17 stable cell line was obtained as described previously (Beaulieu et al., 1996).
Preparation of Microsomal Fraction.
Microsomes were prepared by differential centrifugation. HK293 cells stably expressing UGT2B17 were homogenized in 4 mM K2HPO4 pH 7.0, 20% glycerol, 1 mM EDTA, and 0.5 mM dithiothreitol using a Brinkmann polytron. Cells were centrifuged at 12,000g for 20 min to remove nuclei, unbroken cells, and mitochondria. The supernatant was subsequently centrifuged at 105,000g for 60 min, and the microsomes pellet was resuspended in homogenization buffer and protein contents were determined using the Bradford's reagent, with bovine serum albumin for standard curves. Samples were aliquoted and kept at −80°C until glucuronidation assays.
Glucuronidation Assays.
Initial enzymatic assays were performed using 15 μM of [14C]UDPGA, 100 μM of unlabeled UDPGA, 200 μM of the various aglycons, and 11 μg of microsomal proteins in 50 mM Tris-HCl pH 7.6, 10 mM MgCl2, 100 μg/ml phosphatidylcholine, and 8.5 mM saccharolactone in a final volume of 100 μl for 16 h at 37°C. The assays were terminated by adding 100 μl of methanol and were centrifuged at 14,000g for 1 min to remove precipitated proteins. An aliquot (100 μl) of the aqueous phases were loaded onto thin layer chromatography plates (0.25-mm-thick silica gel 60 F254 S) (EM Scientific, Gibbstown, NJ) and chromatographed in a solvent toluene/methanol/acetic acid (7:3:1). Thin layer chromatography plates were exposed for 1 to 4 days, and the levels of glucuronidation were assessed by PhosphorImager (Amersham Biosciences Inc., Sunnyvale, CA).
The glucuronide formation of compounds that react with UGT2B17 in the screening assay was subsequently quantified in the same buffer containing 15 μM [14C]UDPGA and 500 μM unlabeled UDPGA for 30 min at 37°C. As previously reported (Beaulieu et al., 1996), 500 μM UDPGA and 30 min of incubation are saturating and linear conditions of glucuronidation catalyzed by the UGT2B17 enzyme. Values represent the mean of at least two independent experiments, performed in duplicate.
Km Determination in Microsomal UGT2B17-HK-293 Cells.
Km determinations were performed by incubating UGT2B17-HK293 microsomal proteins with substrates for 30 min at 37°C. Microsomal proteins (15–30 μg) were incubated with 15 μM [14C]UDPGA, 500 μM unlabeled UDPGA, and 5 to 35 μM substrates. The substrates evaluated were alizarin, eugenol, galangin, and scopoletin. Values represent the mean of at least two independent experiments each performed in triplicate.
Results
Screening Assay.
In an initial screening experiment, more than 55 substrates belonging to 11 categories of molecules were incubated in the presence of microsomal preparations from stably transfected UGT2B17-HK293 cells. Whereas several coumarins, nonsteroidal anti-inflammatory drugs (NSAIDs), anthraquinones, flavonoids, monoterpenoids, and phenolic compounds were conjugated, UGT2B17 was unable to glucuronidate the four benzodiazepins and the three opioids tested (Table1). In addition, compounds such as chloramphenicol, ethynylestradiol, hydroxyflutamide, furosemide, 6- and 8-hydroxyquinoline, hecogenin, and α-naphtylamine were not substrates for UGT2B17 (Table 1).
Screening analysis of exogenous compounds glucuronidated by the UGT2B17 enzyme
Coumarins.
This category of molecules contains good substrates for UGT2B17 because 4-methylumbelliferone, umbelliferone, and scopoletin were conjugated (Table 1). In contrast, conjugation was not detected for esculetin and 4-hydroxycoumarin (Table 1). Scopoletin was the best coumarin substrate conjugated by UGT2B17 with a glucuronidation activity of 102.5 pmol · min−1 · mg protein−1 (Fig.1). Although umbelliferone and its 4-methyl derivative were conjugated by UGT2B17, glucuronidation rates were too low to be quantified.
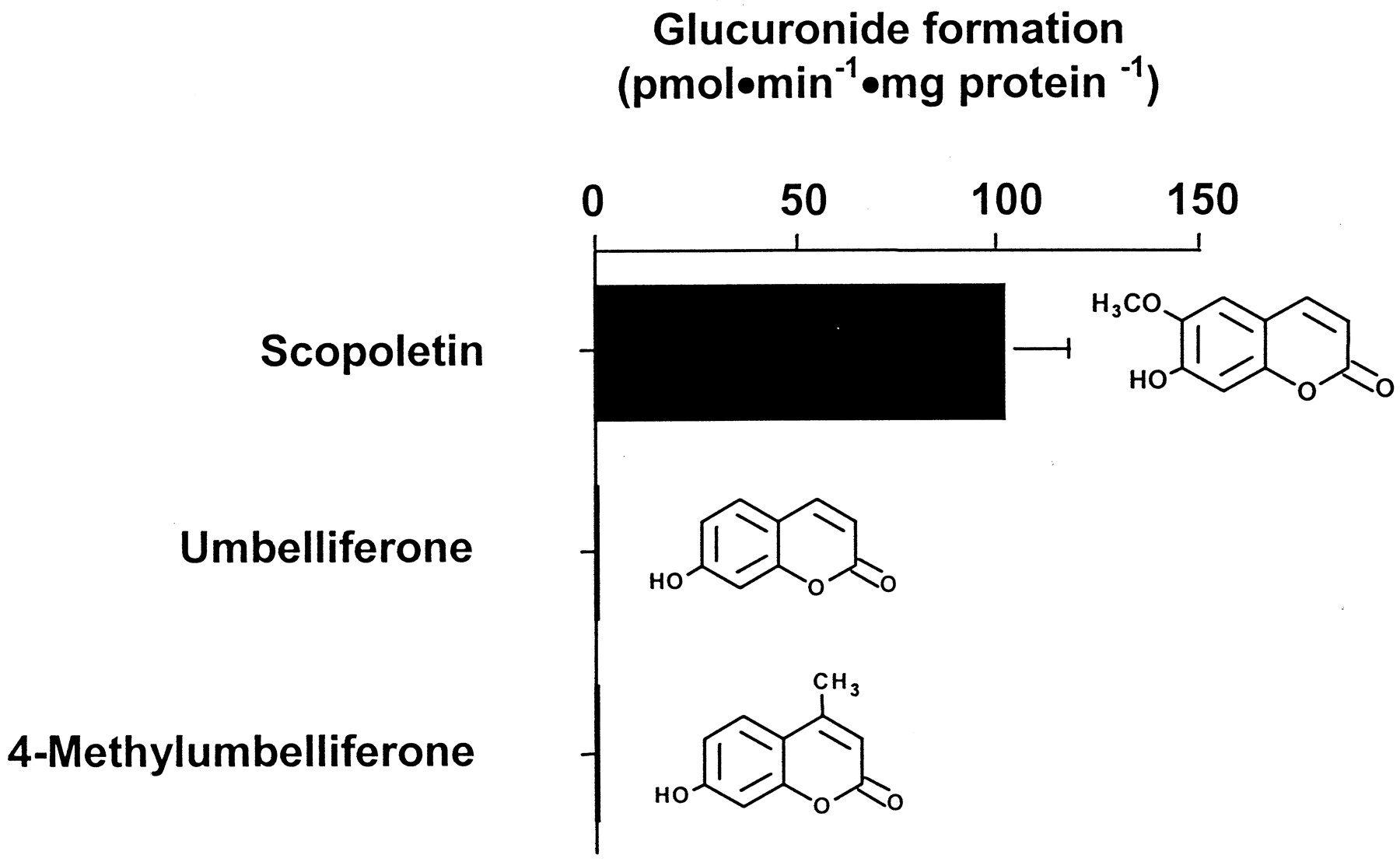
Glucuronidation rates catalyzed by the UGT2B17 enzyme and chemical structure of coumarin derivatives.
Substrates that demonstrated reactivity with UGT2B17 in the screening assay were subsequently reassayed in the presence of 15 μM [14C]UDPGA and 500 μM unlabeled UDPGA for 30 min at 37°C. Vmax values represent the mean of two independent experiments ± S.D.
Anthraquinones.
Anthraquinone molecules are also good substrates for UGT2B17 because only quinizarin was not converted into glucuronide (Table 1). Furthermore, UGT2B17 catalyzed the glucuronidation of alizarin, emodin, and anthraflavic acid with velocities of 125, 43, and 17 pmol · min−1 · mg protein−1, respectively (Fig. 2).

Glucuronidation of anthraquinones by UGT2B17.
Compounds that react with UGT2B17 in the screening were subsequently reassayed as described under Materials and Methods. Glucuronidation values represent the mean of two independent experiments ± S.D.
Flavonoids.
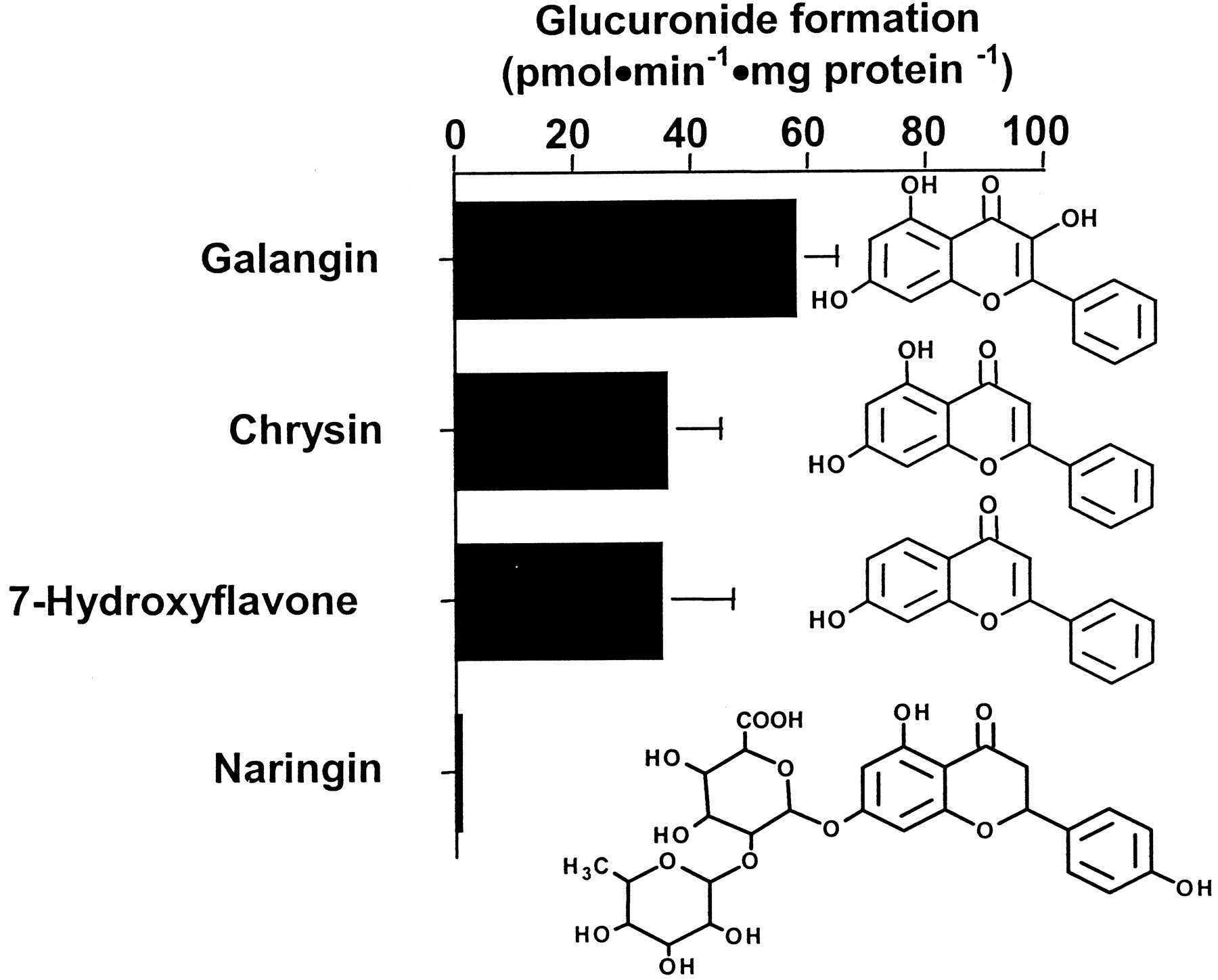
Among flavonoids, galangin, chrysin, 7-hydroxyflavone, and naringin were substrates for the UGT2B17 protein, whereas other flavonoids tested were not conjugated (Table 1). Indeed, glucuronide conversion values of 58, 36, and 35 pmol · min−1 · mg protein−1 were observed for galangin, chrysin, and 7-hydroxyflavone, respectively (Fig.3). The conjugation of naringin was too low to be measured.
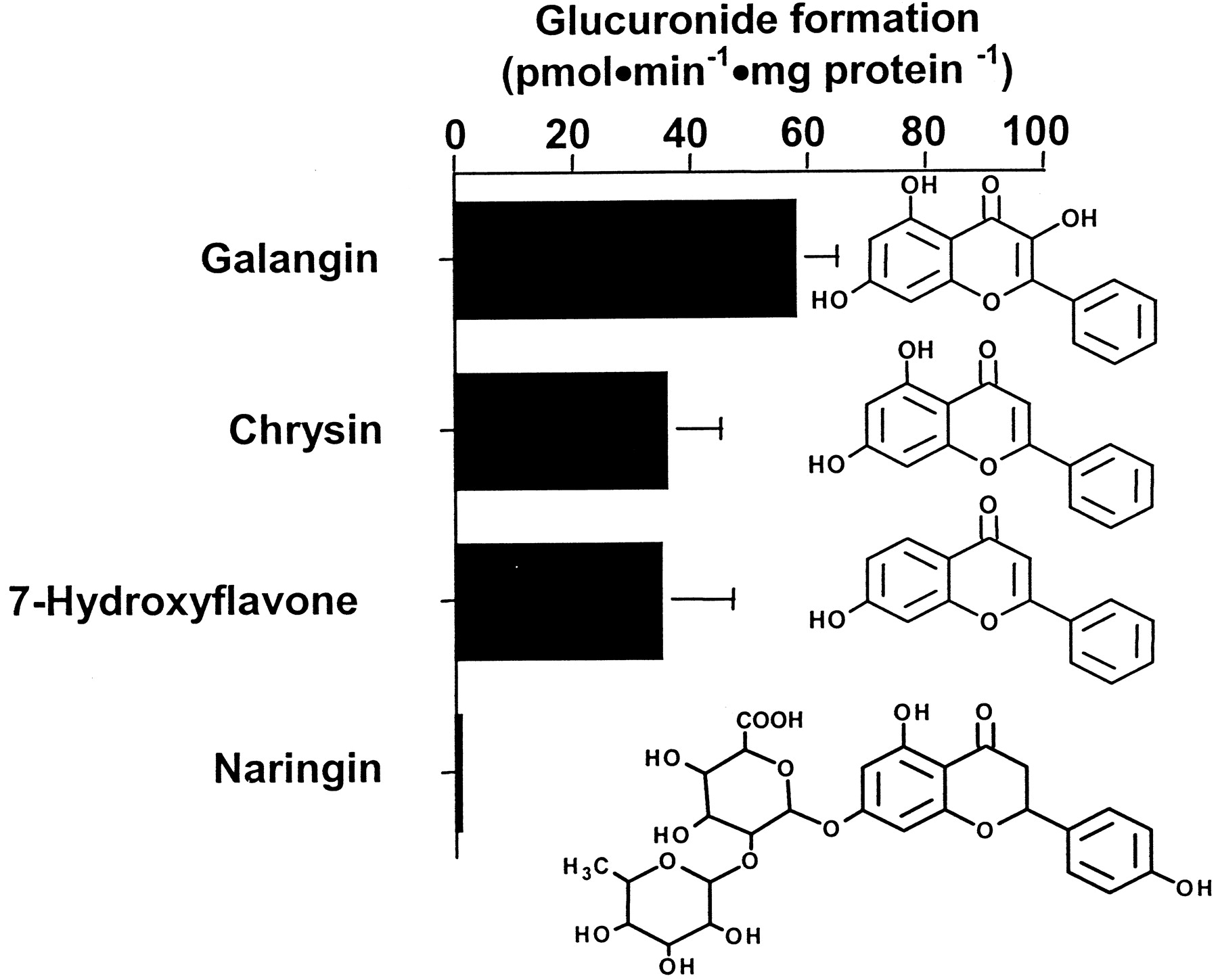
Glucuronidation of flavonoids by the UGT2B17 enzyme and chemical structure of conjugated substrates.
The rate of flavonoid derivatives conjugated by UGT2B17 was determined in the presence of 15 μM [14C]UDPGA and 500 μM unlabeled UDPGA for 30 min at 37°C. Vmaxvalues represent the mean of two independent experiments ± S.D.
NSAIDs and Monoterpenoids.
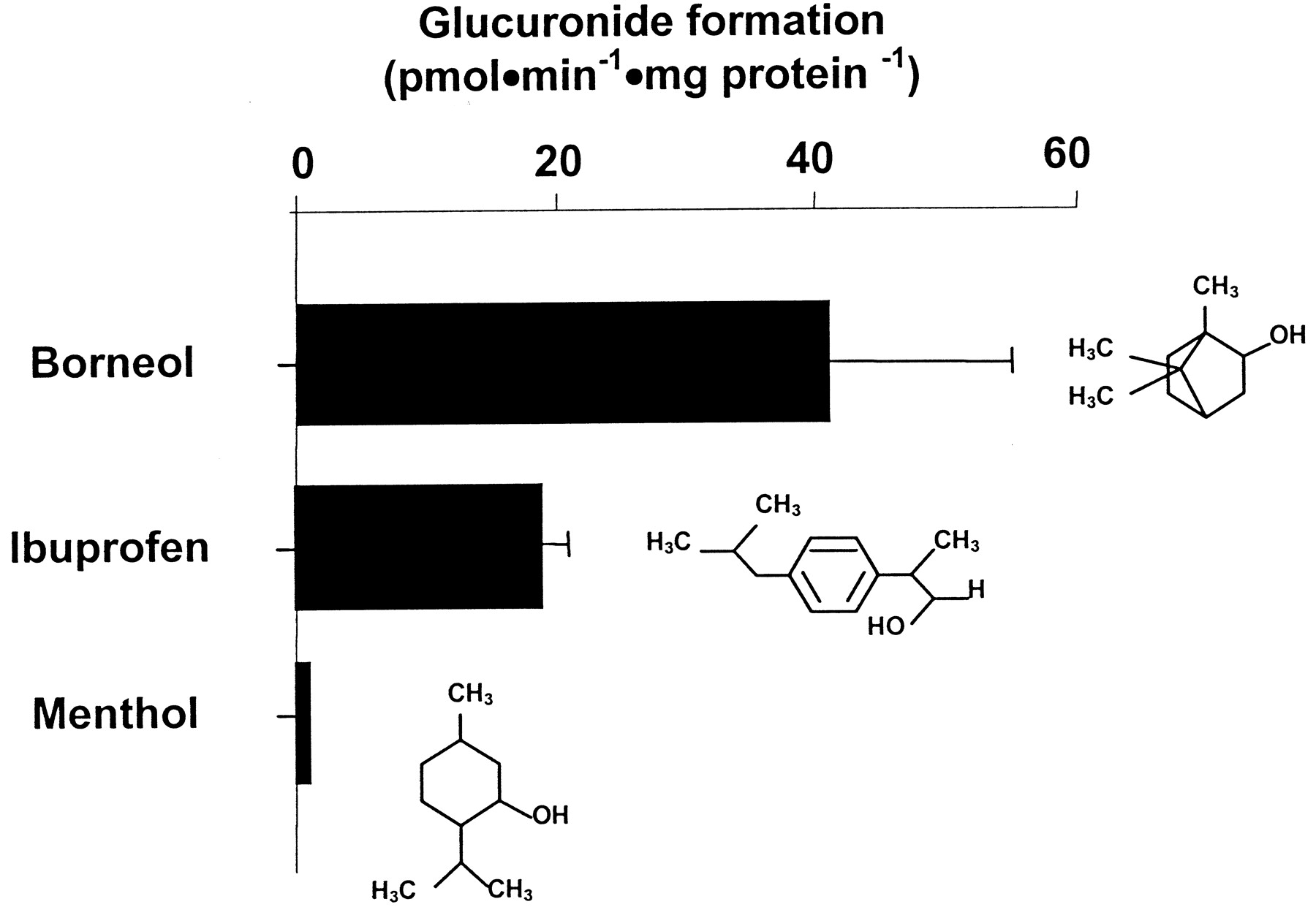
Among the four NSAIDs tested, only ibuprofen was conjugated by the stably expressed UGT2B17 with a conjugation rate of 19 pmol · min−1 · mg protein−1 (Fig.4). Two substrates belonging to the monoterpenoid group of molecules, namely, borneol and menthol, demonstrate glucuronidation in the presence of the UGT2B17 enzyme (Table 1). The glucuronidation rate for borneol was 41 pmol · min−1 · mg protein−1, whereas menthol had a conjugation rate too low to be quantified (Fig. 4).

Glucuronidation rates and chemical structure of borneol, ibuprofen, and menthol derivatives.
Substances that demonstrated reactivity with UGT2B17 in the screening were subsequently quantified. Vmax values represent the mean of two independent experiments ± S.D.
Phenolic Compounds.
Six phenolic compounds were conjugated by UGT2B17: 4-ethyl phenol, 4-propyl phenol, eugenol, 1-naphtol, p-nitrophenol, and phenol red with glucuronidation rates of 43, 32, 102, 36, 32, and 34 pmol · min−1 · mg protein−1, respectively (Fig.5). In contrast, the other phenolic compounds tested such as 4-T-buthylphenol, acetaminophen, caffeic acid, and ferrulic acid were not recognized by UGT2B17 (Table 1).

Glucuronidation rates and chemical structure of phenol derivatives.
Substances that demonstrated reactivity with UGT2B17 in the screening were subsequently quantified. Vmax values represent the mean of two independent experiments ± S.D.
Kinetic Parameters of Glucuronidation.
To further characterize the glucuronidation of exogenous compounds catalyzed by UGT2B17, the kinetic parameters of alizarin, eugenol, galangin, and scopoletin conjugation were determined (Table2). ApparentVmax values varied from 58 to 125.3 pmol · min−1 · mg protein−1 for galangin and alizarin, respectively. The lower apparent Kmvalue was obtained with eugenol (17.9 μM), whereas scopoletin has a low affinity for UGT2B17 with an apparentKm of 78.9 μM. Furthermore, the kinetic analyses revealed that scopoletin has the lowestVmax/Kmratio of the molecules tested with a value of 129.5 l · min−1 · mg−1· 10−8 (Table 2). Alizarin showed the highest glucuronidation efficiency for UGT2B17 with aVmax/Kmratio of 642.6 l · min−1 · mg−1 · 10−8 (Table 2).Vmax/Kmratio of galangin and eugenol were 137.8 and 569.8 l · min−1 · mg−1· 10−8, respectively (Table 2).
Kinetic analysis of exogenous compounds glucuronidation by UGT2B17
Discussion
The expression of the UGT2B17 enzyme in the human prostate, an androgen-sensitive tissue, and its capacity to conjugate androgens have first retained the interest for this enzyme (Beaulieu et al., 1996;Guillemette et al., 1997; Lévesque et al., 1998). However, UGT2B17 is also expressed in the liver, the most important tissue for detoxification. Although the hepatic UGT2B17 should be involved in the clearance of circulating androgens, the implication of other UGT2B enzymes in the hepatic metabolism of xenobiotics suggested a possible role of UGT2B17 in the detoxification of drugs and pollutants to which the human body is exposed. Results from the present study demonstrate that UGT2B17 catalyzes the glucuronide conjugation of several exogenous compounds. Furthermore, it is observed that UGT2B17 conjugates different substrates compared with UGT2B15, another human UGT enzyme that possesses a 95% homology in amino acid sequence with UGT2B17.
More than 55 substrates belonging to 11 major classes of exogenous compounds were screened using the stably transfected UGT2B17-HK293 cell line (Beaulieu et al., 1996). Coumarins, which act as anticoagulants, are mainly found in plants and are commonly used in the treatment of various inflammatory and cerebrovascular diseases (Green et al., 1994;Aherne and O'Brien, 1999). Among this family, scopoletin is the best UGT2B17 substrate having a glucuronidation rate identical to that of eugenol, a UGT substrate commonly used as a positive control for glucuronidation assay. Although the glucuronidation rate of UGT2B17 for scopoletin is relatively high, its glucuronidation efficiency is 4-fold less than that for eugenol. Interestingly, glucuronidation of this inhibitor of the inflammatory response (Silvan et al., 1996) is also catalyzed at a similar conjugation rate by UGT2B15 (Green et al., 1994). In contrast, esculetin is conjugated by UGT2B15 but not by UGT2B17. In addition, glucuronidation rates of UGT2B17 for umbelliferone and 4-methylumbelliferone are very low compared with those of UGT2B15 (Green et al., 1994). Thus, it seems that members of the coumarin family are good substrates for both UGT2B15 and UGT2B17 enzymes and that minor changes in the structure of the UGT2B protein result in significantly different substrate specificities. These data demonstrate that UGT2B15 and UGT2B17 may have distinct and complementary roles in the inactivation of coumarin molecules.
The potential role of NSAIDs in cancer prevention was reported in numerous studies (Schapira et al., 1999). These compounds are also used for pain relief, arthritis, cardiovascular diseases, and Alzheimer. Glucuronidation is the pathway of inactivation and elimination of ibuprofen (Davies, 1998), and previous studies have shown that only UGT2B7, UGT1A3, and UGT1A9, which are expressed in the liver, conjugate ibuprofen (Green and Tephly, 1998; King et al., 2000). In the present study, we demonstrate that this frequently prescribed NSAID is also inactivated by UGT2B17.
Anthraquinones were excellent substrates for UGT2B17 because three of four compounds tested were conjugated. Among this study, alizarin was the substrate with the highest glucuronidation activity and efficiency with UGT2B17. Alizarin is used to prevent recurrence of calcium-containing urinary stones and is eliminated from the body as glucuronide derivatives (Lorenz et al., 1985). Previous studies demonstrated that other UGT enzymes, including UGT1A1, UGT1A3, UGT1A8, UGT1A9, UGT1A10, and UGT2B15 (King et al., 2000) also catalyze alizarin glucuronidation. The same set of enzymes catalyzes the glucuronidation of emodin and anthraflavic acid, with the exception of UGT1A8 and UGT1A10 for which the latter substrate was not tested (King et al., 2000).
Among phenolic compounds, eugenol is the best UGT2B17 substrate, with a glucuronidation rate of 102 pmol · min−1 · mg protein−1. Other phenol substrates tested with UGT2B17 are conjugated with a 2- to 3-fold lower rate than eugenol. In fact, UGT2B17 is active toward simple planar phenols such as 4-ethylphenol, 4-propyl-phenol, 1-naphthol, p-nitrophenol, and phenol red. Thus, our data suggest that the conjugation by this enzyme involves the transfer of glucuronic acid to the phenolic hydroxyl moiety on the aromatic cycle of planar phenols. However, it seems that the presence of a bulky substitute in orthoposition to the hydroxyl group hinders the compound to be glucuronidated by UGT2B17.
Flavonoids are found in vegetables and fruits, in which they provide color, texture, and flavor (Harborne, 1986). They also mediate several biological functions in humans, acting as anticarcinogenic, anti-inflammatory, and antioxidant agents (Aherne and O'Brien, 1999). Flavonoids also regulate drug-metabolizing enzymes, such as P450 cytochromes and glutathione S-transferases expression (Buening et al., 1981) and prevent tumor cell proliferation in the mammary gland (So et al., 1997). The existence of several UGT enzymes able to conjugate flavonoids suggests that glucuronidation is a major metabolic pathway for these compounds (Green et al., 1994; Coffman et al., 1998; King et al., 2000). Interestingly, recent studies demonstrate that galangin was oxidized by P450 cytochromes using liver microsomes but glucuronidation remains the predominant biotransformation pathway (Otake et al., 2002). Of all flavonoids tested, only galangin, chrysin, and 7-hydroxyflavone were conjugated by UGT2B17. These compounds are also conjugated by UGT2B15 (Green et al., 1994). The most striking difference observed between UGT2B15 and UGT2B17 was detected with naringenin glucuronidation, which is catalyzed by UGT2B15 and not by UGT2B17 (Green et al., 1994). This major difference constitutes a useful tool to estimate the activity of the two isoforms within a single tissue.
Related aromatic alcohols, such as menthol, are widely used in a variety of commercial products such as toothpastes, mouthwash, foods, and oral pharmaceutical preparations. Menthol metabolism mainly involves the formation of glucuronide derivatives that are excreted in urine (Gelal et al., 1999). To date, only UGT1A4 and UGT2B7 enzymes were reported as menthol-conjugating enzymes (Coffman et al., 1998). In the present study, it is demonstrated that UGT2B17 also glucuronidates menthol. Other substances commonly used, such as essential oils and sunscreens, contain monoterpenoid components. One of these compounds, borneol, is conjugated by UGT2B17. To date, only UGT1A4 is reported to glucuronidate this compound (King et al., 2000).
Our data demonstrate that UGT2B17 possesses a distinct substrate specificity for xenobiotics. However, some compounds that are glucuronidated by UGT2B17 are also conjugated by other UGT enzymes, indicating that each isoforms may play a redundant role in hepatic detoxification. However, the exact role of each enzyme is also dependent on their relative expression in a given tissue. Recently, the level of several UGT enzymes expression was investigated in the liver from patients with diverse hepatic diseases using specific real-time reverse transcription-polymerase chain reaction analyses (Congiu et al., 2002). Results of this study indicated that UGT2B7, UGT2B15, and UGT2B17 mRNA levels would be approximately in the same range. Because there is a large interindividual variability in the levels of UGT2B mRNA, especially UGT2B17, it is however difficult to compare the expression of UGT2B isoforms in the liver. In addition, this variability also suggests that different UGT enzymes are required for the conjugation of a given exogenous compound in different circumstances or in different individuals.
In summary, the present study clearly demonstrates that UGT2B17 is not an exclusive androgen-conjugating enzyme, because this enzyme glucuronidates substances belonging to different families of xenobiotics. The expression of UGT2B17 in the liver and extrahepatic tissues may contribute to a more efficient elimination of exogenous compounds from the body.
Acknowledgments
We thank Dr. Pei Min Rong for technical assistance. We also thank Dr. Olivier Barbier for critical reading of this manuscript.
Footnotes
-
↵1 These authors contributed equally to this work.
-
This work was supported by the Canadian Institutes of Health Research and the Fonds de la Recherche en Santé du Québec. J.-S.C. is holder of a scholarship from the Canadian Institutes of Health Research and the Canadian Prostate Cancer Research Initiative.
- Abbreviations used are::
- UGT
- UDP-glucuronosyltransferase
- UDPGA
- UDP-glucuronic acid
- HK293
- human embryonic kidney
- NSAID
- nonsteroidal anti-inflammatory drug
- Received November 15, 2002.
- Accepted February 14, 2003.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}