


Abstract
We investigated whether the species difference in the biliary excretion activity of some Mrp2 substrates was attributable to the intrinsic transport potential or the expression level of Mrp2, especially in rat and dog. Dog Mrp2 cDNA was isolated from beagle dog liver, and a vesicle transport study was performed using recombinant rat and dog Mrp2 expressed in insect Sf9 cells. The ATP-dependent transport of 17β-estradiol 17-(β-d-glucuronide) ([3H]E217βG) and leukotriene C4 ([3H]LTC4), normalized by the absolute protein expression level, was similar in both Mrp2s. The Mrp2 protein expression in dog liver was only 10% of that in rat liver and was comparable with the reported difference in the biliary excretion clearance of temocaprilat as Mrp2 substrate. In contrast to LTC4, unique transport kinetics for E217βG were evident in dog Mrp2. In addition to the high-affinity site with a Km value of 3.25 ± 0.10 μM, which is similar to that in rat Mrp2 (4.81 ± 1.21 μM), dog Mrp2 has an additional low-affinity site (≫75 μM), which makes a major contribution to the transport of E217βG (65% of the total transport capacity at tracer concentration). In summary, the difference in the biliary excretion activity of Mrp2 substrates between rat and dog depends on the Mrp2 protein expression level rather than the intrinsic transport activity of the transporter molecules. The unique transport properties of glucuronide conjugates by dog Mrp2 may lead to the species difference involving the drug-drug interaction or drug-induced hyperbilirubinemia on the bile canalicular membrane.
Many endogenous and xenobiotic compounds are excreted into urine and bile via specific transport pathways. In the liver, compounds are taken up into the hepatocytes mainly via specific carrier systems and passive diffusion, metabolized by cytochrome P450 (phase I), conjugated with hydrophilic groups (phase II), and then finally excreted into bile via the efflux transporters (phase III) (Ishikawa, 1992). The multidrug resistance-associated protein 2/ATP-binding cassette transporter family C2 (MRP2/ABCC2) is the major primary active efflux transporter involved in the phase III biliary excretion process of many organic anions (Suzuki and Sugiyama, 1998; Keppler and König, 2000). Mrp2 substrates include glutathione, glutathione conjugates, glucuronide conjugates, sulfated and glucuronidated bile salts, and nonconjugated organic anions (Suzuki and Sugiyama, 1998; Keppler and König, 2000). Mrp2 accounts for most of the bile salt-independent flow (BSIDF). The physiological significance of Mrp2 was reconfirmed by the finding of some nonsense and missense mutations in MRP2 from patients with Dubin-Johnson syndrome and in animal models (TR- rats and Eisai hyperbilirubinemic rats) that exhibit hyperbilirubinemia (Paulusma and Oude Elferink, 1997; König et al., 1999; Suzuki and Sugiyama, 2002). The Mrp2 cDNA sequence has already been reported in various mammalian species, including human (4635 bp) (Taniguchi et al., 1996; Paulusma et al., 1997), rat (4623 bp) (Buchler et al., 1996; Paulusma et al., 1996; Ito et al., 1997), mouse (4629 bp) (Fritz et al., 2000), monkey (4632 bp), rabbit (4692 bp) (van Kuijck et al., 1996), and dog (4506 and 4632 bp) (Conrad et al., 2001); however, functional analysis of the cloned Mrp2 cDNA product has not been reported except in human, rat, and rabbit (Madon et al., 1997; Ito et al., 1998; van Aubel et al., 1998; Cui et al., 1999). Several reports suggested a large species difference in the biliary excretion clearance of organic anions, especially Mrp2 substrates. Niinuma et al. (1999) reported that the ATP-dependent transport of glutathione conjugates and nonconjugated organic anions into bile canalicular membrane vesicles (CMVs) from human liver was lower than that into CMVs from rat liver, although the ATP-dependent transport of glucuronide conjugates was similar in human and rat (Niinuma et al., 1999). Ishizuka et al. (1997) also reported a large species difference in the biliary excretion clearance of temocaprilat, an Mrp2 substrate (Ishizuka et al., 1997), in vivo and also in the ATP-dependent transport activity of 2,4-dinitrophenyl-S-glutathione (DNP-SG) into CMVs in vitro (rat ≫ mouse > guinea pig > rabbit > dog) (Ishizuka et al., 1999). During the drug development process, species differences in the pharmaco- and toxicokinetics of candidate drugs are a major problem. Possibly, Mrp2 transport activity, as well as inhibitor sensitivity, could lead to various phenomena (e.g., the difference in the biliary excretion clearance, the delay in the elimination half-life from the circulating blood, and the toxic side effects on the Mrp2-mediated drug-drug interaction, such as drug-induced hyperbilirubinemia). Despite such species difference in the biliary excretion potential of Mrp2 substrates, the precise mechanism (i.e., difference in the protein expression level and/or intrinsic transport properties of a single transporter molecule and/or transport affinity) has not yet been elucidated.
In the present study, we investigated the transport characteristics of cloned dog Mrp2 in comparison with rat Mrp2 using respective standard antigens to normalize the absolute protein expression level. Moreover, Mrp2 protein expression levels in the livers were examined in an attempt to identify the cause of the previously reported large species difference in the biliary excretion of Mrp2 substrates between rat and dog.
Materials and Methods
Materials. [3H]17β-Estradiol 17-(β-d-glucuronide) (E217βG; 55 Ci/mmol, 97%) and [3H]leukotriene C4 (LTC4, 100 Ci/mmol, 95%) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). E217βG and 4-methylumbelliferyl-β-d-glucuronide (4-MUG) were purchased from Sigma-Aldrich (St Louis, MO). LTC4 was purchased from Cayman Chemical (Ann Arbor, MI). All other chemicals were of analytical grade. Sf9 cells were maintained as a suspension culture at 27°C with serum-free EX-CELL 420 medium (JRH Biosciences, Inc., Lenexa, KS). The beagle dog liver was kindly donated by Sankyo Co., Ltd. (Tokyo, Japan).
Plasmid Construction. To obtain dog Mrp2 cDNA, polymerase chain reaction (PCR) was performed using the forward primer [5′-aaacccAGCGCTgccatgctggagaagttctgcaactctacgttttgg-3′, which includes the Aor51HI site (single underline)] and the reverse primer [5′-cagggaAAGCTTtcagaacgatgtgctgttcacattttcaatgcc-3′, which includes the HindIII site (double underline)] with beagle dog liver cDNA by KOD-plus DNA polymerase (Toyobo Co., Ltd., Osaka, Japan). A PCR product of approximately 4.6 kilobases was subsequently digested at the HindIII site located in the reverse primer sequence. This fragment was inserted into pBluescript II SK(-) (Stratagene, La Jolla, CA), digested with SmaI-HindIII in the multiple cloning site of the vector, and its sequence was analyzed. To construct dog Mrp2 cDNA containing baculovirus-expression vector, the cDNA cassette was digested with Aor51HI and HindIII and inserted into the SmaI-HindIII site of recombinant donor plasmid after removing the rat Mrp2 cDNA cassette from the previously reported rat Mrp2-pFASTBAC1 using the same enzyme combination (Ito et al., 2001b). Homologous recombination in Escherichia coli DH10BAC was performed according to the manufacturer’s instructions to obtain the recombinant bacmid.
Production and Infection of Recombinant Baculovirus and Membrane Vesicle Preparation. Recombinant baculoviruses carrying dog Mrp2 or rat Mrp2 were prepared as described with some modifications (Ito et al., 2001b). Briefly, Sf9 cells were transformed with the bacmid carrying dog or rat Mrp2 cDNA using a FuGENE 6 transfection reagent (Roche Diagnostics, Indianapolis, IN). After more than three rounds of amplification, a high-titer virus supernatant was obtained. The Sf9 cell suspension was poured into a culture dish and stood for 1 h at 27°C. During this incubation, the cells were allowed to attach to the dish. The medium was changed to fresh medium supplemented with 5% fetal bovine serum and recombinant baculoviruses. In our experiments, the multiplicity of infection was 1 to 10. For control experiments, Sf9 cells were infected with a baculovirus encoding the green fluorescent protein (GFP). Cells were harvested 60 h after infection, and subsequently, membrane vesicles were isolated from 1 to 2 × 108 Sf9 cells using a standard method described previously with some modifications (Müller et al., 1994; Ito et al., 2001b). Briefly, cells were diluted 40-fold with hypotonic buffer (1 mM Tris-HCl and 0.1 mM EDTA, pH 7.4 at 4°C) containing 2 mM phenylmethylsulfonyl fluoride, 5 μg/ml leupeptin, 1 μg/ml pepstatin, and 5 μg/ml aprotinin and stirred gently for 1 h on ice. The cell lysate was centrifuged at 100,000g for 30 min at 4°C, and the resulting pellet was suspended in 10 ml of isotonic TS buffer (10 mM Tris-HCl and 250 mM sucrose, pH 7.4 at 4°C) and homogenized in a Dounce B homogenizer (glass/glass, tight pestle, 30 strokes). The crude membrane fraction was layered on the top of a 38% (w/v) sucrose solution in 5 mM Tris-HEPES, pH 7.4 at 4°C, and centrifuged in a Hitachi P40ST rotor at 280,000g for 45 min at 4°C. The turbid layer at the interface was collected, diluted to 23 ml with TS buffer, and centrifuged at 100,000g for 30 min at 4°C. The resulting pellet was suspended in 400 μl of TS buffer. Vesicles were formed by passing the suspension 30 times through a 25 gauge needle with a syringe. The membrane vesicles were finally frozen in liquid nitrogen and stored at -80°C until use. The protein concentration was determined by the Lowry method (Lowry et al., 1951) using bovine serum albumin (BSA) as a standard.
Production of Standard Antigen for Human, Rat, and Dog Mrp2. The cDNA fragments encoding the carboxy-terminal 53 amino acids of dog, rat, and human Mrp2 were amplified from mRNA of rat, dog, and human liver using the universal forward primer [5′-cacaggGAATTCaccatcatggacagtgac-3′ with an EcoRI linker (single underline)] and respective reverse primer [5′-ccGTCGACtagaattttgtgctgttc-3′ for human, 5′-ccGTCGACtagagctctgtgtgatt-3′ for rat, and 5′-gggacGTCGACtcagaacgatgtgctgttc-3′ for dog with a SalI site (double underline)]. PCR products of approximately 170 bases were subsequently digested with EcoRI-SalI and then inserted into the EcoRI-SalI site of pMAL-c2 vector (New England Biolabs, Beverly, MA). These vectors were used to transform E. coli-competent DH5α. After induction with 0.3 mM isopropyl-β-d-galactopyranoside for 2 h at 37°C, total E. coli lysates were subjected to SDS-polyacrylamide gel electrophoresis followed by Coomassie Brilliant Blue staining to quantify the amount of protein by densitometric analysis in comparison with BSA as a standard.
Isolation of Crude Membrane from the Liver. The crude membrane was isolated from liver as described previously (Vos et al., 1998). The liver was cut into small pieces (about 1 mm) with scissors and homogenized in buffer A (1 mM NaHCO3, 0.5 mM phenylmethylsulfonyl fluoride, 5 μg/ml leupeptin, 1 μg/ml pepstatin, and 5 μg/ml aprotinin) with a Dounce homogenizer (glass/glass, loose pestle, 20 strokes). The homogenate was diluted 6-fold with buffer B (1 mM NaHCO3 and 0.5 mM phenylmethylslfonyl fluoride) and centrifuged at 1500g for 15 min at 4°C. The pellet was resuspended in a 2.5-fold volume of 70% (w/w) sucrose and overlaid with 44% sucrose (w/w; density = 1.1972) and 36.5% sucrose (w/w; density = 1.1587), and centrifuged in a Hitachi P40ST rotor at 100,000g for 90 min at 4°C. The layer of the interface between the 36.5 and 44% sucrose was collected and suspended in buffer B. The sample was centrifuged at 100,000g for 20 min at 4°C, and then the pellet was resuspended in buffer A. Samples were homogenized by passing them 15 times through a 25 gauge needle and stored at -80°C until Western blot analysis.
Western Blot Analysis. Membrane vesicles from Sf9 cells and crude membranes from rat and dog liver were loaded onto an 8.5% polyacrylamide slab gel containing 0.1% SDS and then transferred to an Immobilon-P Transfer Membrane filter (Millipore Corporation, Bedford, MA) by electroblotting. The filter was blocked at room temperature for 1 h with TTBS (Tris-buffered saline with 0.05% Tween 20) containing 3% BSA and probed at room temperature for 1 h with primary antisera diluted with TTBS containing 0.1% BSA. Used antisera were rabbit anti-rat Mrp2 antiserum and anti-human MRP2 antiserum raised against the carboxy-terminal amino acid residues of 1530 to 1541 for rat and 1534 to 1545 for human, respectively, as reported previously (Büchler et al., 1996; Schaub et al., 1999). The filters were incubated at room temperature for 1 h with donkey anti-rabbit IgG conjugated with horseradish peroxidase (Santa Cruz Biotechnology, Santa Cruz, CA) diluted with TTBS containing 0.1% BSA (1:3000) and examined using the enhanced chemiluminescence detection kit (Amersham Biosciences, Piscataway, NJ). Images were analyzed using an LAS1000 chemiluminescence detector (Fuji Photo Film Co. Ltd., Tokyo, Japan). The Mrp2 expression level was calculated from the band density using rat and dog Mrp2 carboxy-terminal fusion protein as standards. Serial dilution and the exposure time were chosen so that the densities were in the linear range.
Transport Study. The vesicle transport study was performed using the rapid filtration technique described previously (Ito et al., 1998). Briefly, 16 μl of transport medium (10 mM Tris, 250 mM sucrose, 10 mM MgCl2, 5 mM ATP, or AMP, pH 7.4 at 37°C) containing radiolabeled compounds (50 nM [3H]E217βG and 5 nM [3H]LTC4) was preincubated for 3 min at 37°C ([3H]E217βG) or at 25°C ([3H]LTC4) and then rapidly mixed with 4 μl of membrane vesicle suspension (5–10 μg of protein). The transport reaction was stopped by the addition of 1-ml ice-cold buffer containing 250 mM sucrose, 100 mM NaCl, and 10 mM Tris-HCl (pH 7.4). The stopped reaction mixture was filtered through a 0.45-μm HAWP filter (Millipore Corporation) and then washed twice with 5-ml stop buffer. The radioactivity retained on the filter was determined using a liquid scintillation counter (LSC5000, Aloka Co., Tokyo, Japan). The initial transport of a tracer amount of each ligand was determined in the presence of AMP or ATP; [3H]E217βG (50 nM at 30 s), [3H]LTC4 (5 nM at 30 s). The Mrp2-dependent transport of [3H]E217βG or [3H]LTC4 was calculated by subtracting the transport into GFP-control vesicles from that into Mrp2-expressing vesicles in the presence of ATP. The intrinsic transport activities of Mrp2 were obtained by dividing the Mrp2-dependent transport of these ligands (pmol/min/mg protein) by the absolute Mrp2 expression level (pmol Mrp2 molecules/mg protein). Kinetic parameters for the transport of [3H]LTC4 were estimated from the following equation:  where v is the transport rate of [3H]LTC4, Vmax is the maximum transport rate (pmol/min/mg protein), S is the ligand concentration in the medium (μM), n is a Hill coefficient, and Km is the Michaelis constant (μM). On the other hand, kinetic parameters for the transport of [3H]E217βG were estimated from the following equation:
where v is the transport rate of [3H]LTC4, Vmax is the maximum transport rate (pmol/min/mg protein), S is the ligand concentration in the medium (μM), n is a Hill coefficient, and Km is the Michaelis constant (μM). On the other hand, kinetic parameters for the transport of [3H]E217βG were estimated from the following equation:  where PSns is the nonsaturable component (μl/min/mg protein). The kinetic data were fitted to the above equations by an iterative nonlinear least-squares method using the MULTI program to obtain kinetic parameters (Yamaoka et al., 1981). The input data were weighted as the reciprocal of the square of the observed values, and the algorithm used for the fitting was the Damping Gauss-Newton Method.
where PSns is the nonsaturable component (μl/min/mg protein). The kinetic data were fitted to the above equations by an iterative nonlinear least-squares method using the MULTI program to obtain kinetic parameters (Yamaoka et al., 1981). The input data were weighted as the reciprocal of the square of the observed values, and the algorithm used for the fitting was the Damping Gauss-Newton Method.
Determination of Inhibition Parameters. The inhibition constants of 4-MUG for the Mrp2-dependent transport of [3H]E217βG were calculated by fitting the data into the following equations: 
 where R is the inhibition-ratio, I is the inhibitor (4-MUG) concentration in the medium (μM), IC50 is the inhibitor concentration at a half-inhibition effect, and r is contribution-ratio of the higher affinity component. The inhibition-ratio data were fitted to the above equations by an iterative nonlinear least-squares method using the MULTI program to obtain inhibition parameters (Yamaoka et al., 1981). The input data were weighted as the reciprocal of the square of the observed values, and the algorithm used for the fitting was the Damping Gauss-Newton Method. The consistency of whether there was a single component (eq. 1) or two components (eq. 2) was evaluated based on the Akaike’s information criterion (AIC) value. The AIC value was used to statistically determine the number of parameters in an equation that adequately represents a group of experimental data. Theoretically, the equation with the minimum AIC is regarded as the best representation of the experimental data (Yamaoka et al., 1978).
where R is the inhibition-ratio, I is the inhibitor (4-MUG) concentration in the medium (μM), IC50 is the inhibitor concentration at a half-inhibition effect, and r is contribution-ratio of the higher affinity component. The inhibition-ratio data were fitted to the above equations by an iterative nonlinear least-squares method using the MULTI program to obtain inhibition parameters (Yamaoka et al., 1981). The input data were weighted as the reciprocal of the square of the observed values, and the algorithm used for the fitting was the Damping Gauss-Newton Method. The consistency of whether there was a single component (eq. 1) or two components (eq. 2) was evaluated based on the Akaike’s information criterion (AIC) value. The AIC value was used to statistically determine the number of parameters in an equation that adequately represents a group of experimental data. Theoretically, the equation with the minimum AIC is regarded as the best representation of the experimental data (Yamaoka et al., 1978).
Results
Isolation of Dog Mrp2 from Beagle Dog Liver. Cloned dog Mrp2 cDNA consisted of a 4632-bp open reading frame (ORF) that encodes the 1544 predicted amino acid sequence. It contained 10 nucleotide substitutions which led to seven amino acids mutations compared with the reported dog Mrp2 sequence (Genbank accession AJ315401) (Fig. 1). Those 10 nucleotide substitutions were also identified in direct sequencing analysis of the partial dog Mrp2 fragments (approx. 500 bp each) covering the entire ORF region (data not shown); therefore, these substitutions were considered as single nucleotide polymorphisms in dog Mrp2. Among those seven amino acid mutations, V1109I was in the predicted transmembrane (TM) 14 region, and the other amino acid mutations (E660D, L665Q, M666L, A861T, G1403E, and R1479K) were in the predicted cytoplasmic region (Fig. 1).

Comparison of our dog Mrp2 with the reported sequence. A, schematic diagram of the distribution of amino acid substitutions in dog Mrp2. Dots represent the approximate location of amino acid substitutions compared with the reported dog Mrp2 sequence (Genbank accession No. AJ315401). NBD, nucleotide binding domain. B, detailed alignment of the amino acid substitutions in dog Mrp2. a, amino acid sequence of dog Mrp2 reported previously (AJ315401). b, amino acid sequence of dog Mrp2 obtained in this study.
Quantification of Rat and Dog Mrp2 Protein Expression in the Membrane Vesicles. We tried to detect rat and dog Mrp2 protein in a quantitative manner using two kinds of antisera against rat and human Mrp2/MRP2 because we had no specific antiserum against dog Mrp2. Anti-rat Mrp2 and anti-human MRP2 antisera have been prepared in rabbits by immunizing the c-terminal 12 amino acid peptide of rat Mrp2 and human MRP2, respectively (Büchler et al., 1996; Schaub et al., 1999). Since the Mrp2 c-terminal amino acid sequences of human, rat, and dog share a high identity (Fig. 2A), it is very likely that the dog Mrp2 c-terminal also cross-reacts with anti-rat Mrp2 and/or anti-human MRP2 antiserum. We checked the reactivity of these two antisera against human, rat, and dog Mrp2 using standard Mrp2 peptides (c-terminal 53 amino acids) fused with MBP. As shown, dog Mrp2 c-terminal MBP fusion protein was detected with anti-human MRP2 antiserum with an efficiency of 33.3% compared with that of human MRP2 c-terminal MBP fusion protein (Fig. 2B). On the other hand, anti-rat Mrp2 antiserum scarcely reacted with dog Mrp2 c-terminal MBP fusion protein (efficiency of less than 1% compared with rat Mrp2 c-terminal MBP fusion protein) (Fig. 2C). Based on these observations, anti-rat Mrp2 antiserum and anti-human MRP2 antiserum were used to detect rat Mrp2 and dog Mrp2, respectively.

Preparation of standard MBP fusion proteins of each Mrp2. A, detailed alignment of human, rat, and dog Mrp2 c-terminal region (53 amino acid residues) is shown. These Mrp2 c-terminal regions were fused with MBP to produce standard Mrp2 proteins as described under Materials and Methods. B, 100 fmol of each standard Mrp2 protein was subjected to Western blot analysis and detected with anti-human MRP2 antiserum (left panel) and anti-rat Mrp2 antiserum (right panel). C, the relative reactivity (%) of anti-human MRP2 antiserum and anti-rat Mrp2 antiserum against the standard Mrp2 proteins.
To quantify the absolute Mrp2 expression level in membrane vesicles from rat and dog Mrp2-expressing Sf9 cells, quantitative Western blot analysis was performed using the respective standard MBP fusion proteins (Fig. 3). Single immunoreactive bands with similar molecular weight were observed in both vesicles (175 kDa). The calculated expressions were 524 ± 55 and 409 ± 21 pmol Mrp2/mg protein for rat and dog Mrp2, respectively.

Quantitative analysis of Mrp2 protein expression in Sf9 membrane vesicles from rat and dog Mrp2-expressing Sf9 cells. The membrane vesicles (0.5 μg of protein) and respective standard Mrp2 protein (50–400 fmol) were separated on an 8.5% polyacrylamide slab gel containing 0.1% SDS. The fractionated proteins were transferred to a membrane filter by electroblotting. Rat and dog Mrp2 were detected with anti-rat Mrp2 (A) and anti-human MRP2 (B) antiserum, respectively. The absolute expressions of rat and dog Mrp2 were calculated from the standard curve of each standard Mrp2 protein. The band densities of Mrp2s in Sf9 cells were within a linear range of the respective standard Mrp2 proteins.
Transport of Typical Mrp2 Substrates by Rat and Dog Mrp2. A transport experiment was performed with rat and dog Mrp2-expressing membrane vesicles as used in Fig. 3. ATP-dependent transport of [3H]LTC4 and [3H]E217βG was readily observed in both vesicles (Fig. 4), confirming that dog Mrp2 accepts glutathione and glucuronide conjugates as substrates. ATP-dependent transport of these substrates was not observed in GFP-expressing vesicles (Fig. 4). Transport in the absence of ATP (i.e., in the presence of AMP) was negligible (0.22 ± 0.11 pmol/mg protein/2 min for [3H]LTC4 and 0.11 ± 0.24 pmol/mg protein/2 min for [3H]E217βG) and also not affected by the expression of rat and dog Mrp2 (data not shown). The initial rates of the ATP-dependent transport of [3H]LTC4 (30 s) and [3H]E217βG (30 s) into dog Mrp2 expressing vesicles were 3.25 ± 0.22 and 1.46 ± 0.36 pmol/mg protein/30 s, respectively. To compare the intrinsic transport activity of rat and dog Mrp2, the initial rates of ATP-dependent transport were further normalized by the absolute expression level of rat and dog Mrp2 protein as determined above. Finally, the transport activities of rat Mrp2 for [3H]LTC4 and [3H]E217βG were 3.50 ± 0.40 and 4.26 ± 0.97 pmol/nmol Mrp2/30 s, respectively, and those of dog Mrp2 for [3H]LTC4 and [3H]E217βG were 7.94 ± 0.54 and 3.57 ± 0.87 pmol/nmol Mrp2/30 s, respectively. The intrinsic transport activity of dog Mrp2 is somewhat higher for [3H]LTC4 and similar for [3H]E217βG compared with that of rat Mrp2.
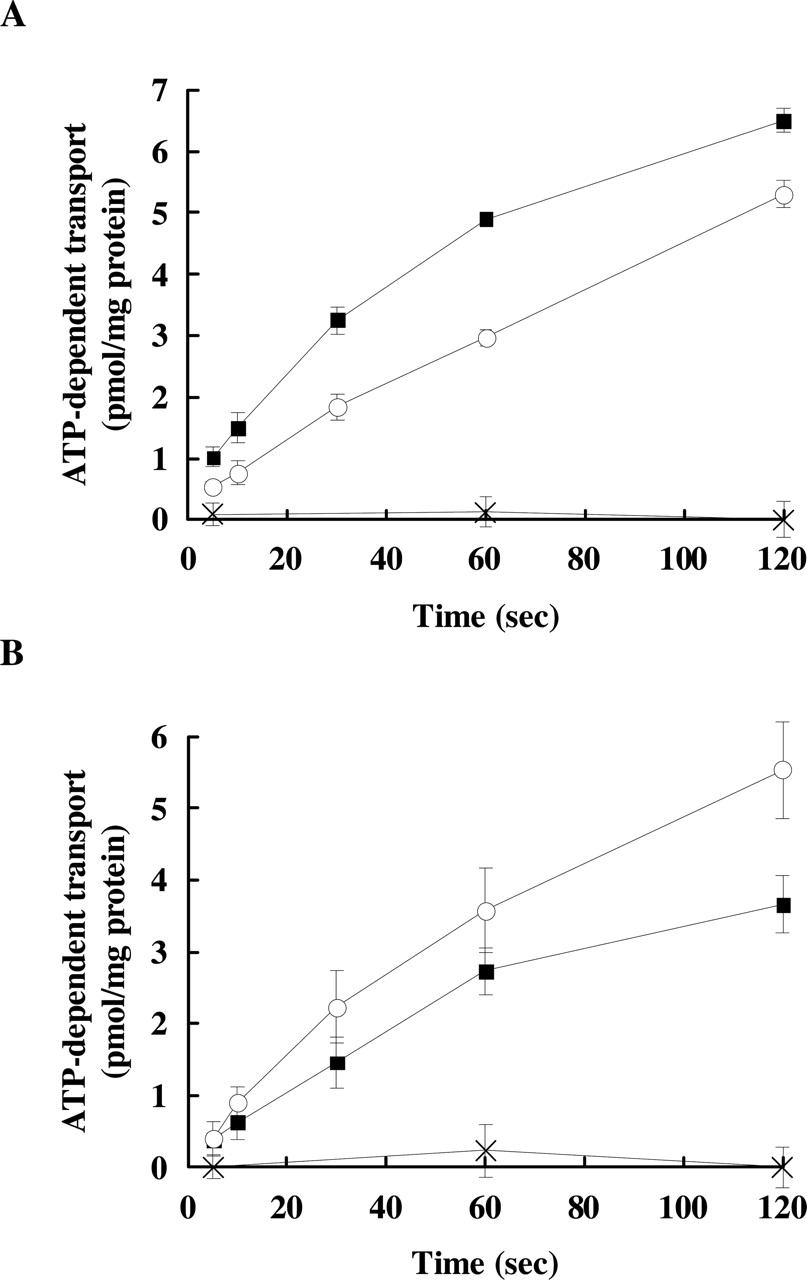
The ATP-dependent transport of typical Mrp2 substrates by rat and dog Mrp2. The membrane vesicles (5–10 μg of protein) from respective rat Mrp2- (○), dog Mrp2- (▪), and GFP- (×) expressing Sf9 cells were incubated in medium containing 5 nM [3H]LTC4 (A) at 25°C or 50 nM [3H]E217βG (B) at 37°C in the presence of 5 mM ATP or AMP. Results are shown as the ATP-dependent transport calculated by subtracting the transport in the presence of AMP from that in the presence of ATP. Each point and vertical bar represents the mean ± S.D. of triplicate determinations.
Osmolarity-Dependent Transport of [3H]E217βG into the Sf9 Membrane. To confirm that vesicle-associated transport of [3H]E217βG reflects transport into a intravesicular space rather than binding to the vesicle surface, the osmolarity-dependence of [3H]E217βG transport was investigated. The initial rates of [3H]E217βG transport into rat and dog Mrp2-expressing membrane vesicles in the presence of ATP were reduced linearly with increasing the extravesicular sucrose concentration from 250 to 1000 mM (Fig. 5). Apparently, surface binding in the presence of ATP was negligible, as the transport was virtually diminished at higher osmolarity condition. Both the transport and surface binding were also negligible in the presence of AMP.

Osmolarity dependence of [3H]E217βG transport into membrane vesicles. The membrane vesicles (5–10 μg of protein) from rat Mrp2- (○, •), dog Mrp2- (□, ▪), and GFP- (×, *) expressing Sf9 cells were incubated at 37°C for 30 s in medium containing 50 nM [3H]E217βG at various concentrations of sucrose (0.25, 0.33, 0.50, and 1.0 M) in the presence of 5 mM ATP (•, ▪, *) or AMP (○, □, ×), respectively. Each point and vertical bar represents the mean ± S.D. of triplicate determinations.
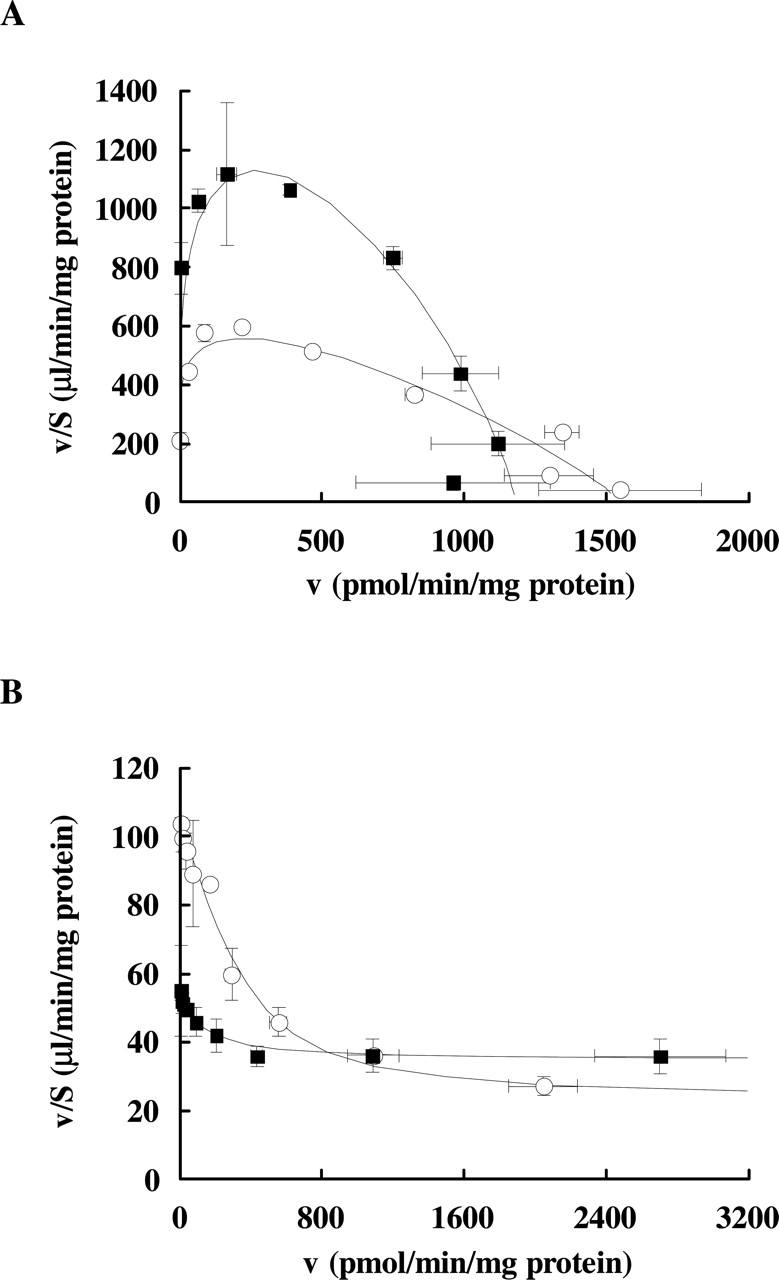
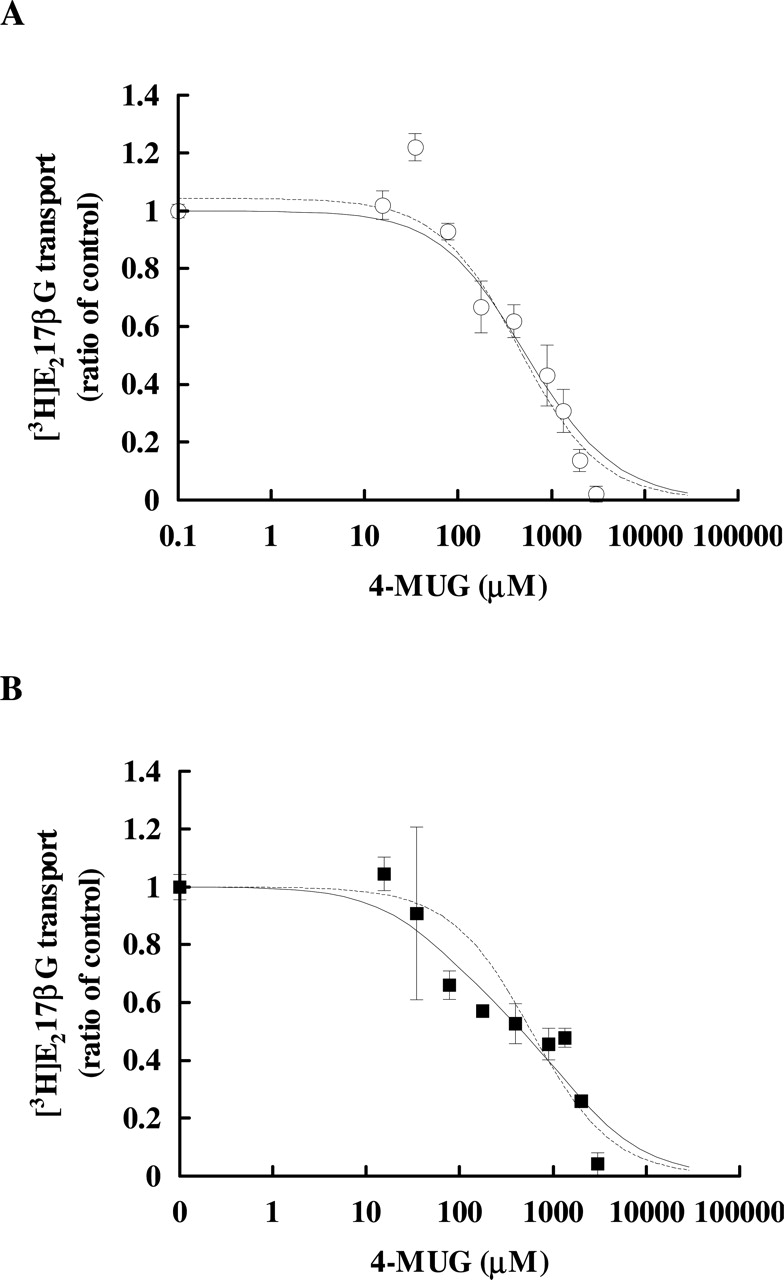
Concentration-Dependence of [3H]LTC4 and [3H]E217βG Transport. To investigate the differences in the affinity of glutathione and glucuronide conjugates between rat and dog Mrp2, the concentration dependence in the transport of [3H]LTC4 and [3H]E217βG was examined (Fig. 6). The initial rates of the Mrp2-dependent transport were determined at several concentrations of LTC4 (0.004–35 μM) and E217βG (0.1–75 μM). The kinetic parameters before and after normalization with the respective Mrp2 expression level are shown in Table 1. The concentration-dependent transport of LTC4 in both rat and dog Mrp2-expressing vesicles exhibited allosteric nature fitting to the Michaelis-Menten equation with a Hill coefficient (rat, 1.16 and dog, 1.30; respectively). The calculated Km values of [3H]LTC4 were comparable between rat and dog; 2.05 ± 0.41 μM and 0.53 ± 0.03 μM for rat and dog Mrp2, respectively (Fig. 6A, Table 1). On the other hand, there was a marked difference in the concentration-dependence of [3H]E217βG transport (Fig. 6B, Table 1). The Mrp2-dependent transport of E217βG consisted of two sites in both rat and dog Mrp2-expressing vesicles, a similar high-affinity site (Km of 4.81 ± 1.21 and 3.25 ± 0.10 μM for rat and dog Mrp2, respectively, Table 1) and lower nonsaturable sites (PSns of 22.7 ± 0.41 and 34.7 ± 0.23 μl/min/mg protein for rat and dog Mrp2, respectively, Table 1). Notably, the contribution ratio of these two transport sites was quite different between rat and dog Mrp2. Under tracer conditions (50 nM), the contribution of the high-affinity saturable site (calculated from Vmax/Km) was as high as 80%, and the lower nonsaturable site was about 20% of the total transport in rat Mrp2 (Table 1). On the other hand, the contribution of the latter low-affinity site was even higher (65%) than the high-affinity saturable site (35%) in dog Mrp2 (Table 1). Supporting the existence of such a low-affinity site for glucuronide conjugate transport, an inhibition study was performed with 4-MUG on the transport of 50 nM [3H]E217βG (Fig. 7 and Table 2). The transport of [3H]E217βG in rat and dog Mrp2 was wholly inhibited by 3 mM 4-MUG (Fig. 7). The inhibition profile in dog Mrp2 showed two components rather than a single component. IC50 values of these two components were calculated to be 70.1 ± 51.8 μM and 1.54 ± 0.16 mM, respectively. The contributions of the higher and lower affinity sites were calculated to be 41.8 ± 15.3 and 58.2 ± 15.3% of the total transport, respectively. In rat Mrp2, the inhibition profile was indistinguishable between the single site model and the two-site model.

Concentration dependence of LTC4 and E217βG. Transport of [3H]LTC4 for 30 s at 25°C (A) or [3H]E217βG for 30 s at 37°C (B) was examined in the presence of various concentrations of substrates for rat Mrp2 (○) and dog Mrp2 (▪). Each symbol represents the Mrp2-dependent transport, determined by subtracting the transport into GFP-control vesicles from that into Mrp2-expressing vesicles in the presence of ATP. Each point and vertical/horizontal bar represents the mean ± S.D. of triplicate determinations. Data were fitted to respective Michaelis-Menten equation as described under Materials and Methods.
Kinetic parameters for the transport of LTC4 and E217ßG by rat and dog Mrp2
Each value was calculated from the data shown in Fig. 6. Values represent the obtained parameter ± computer calculated S.D. V′max and PS′ns values mean Vmax and PSns values normalized by the absolute expression amount of each Mrp2 protein, respectively.

Inhibitory effect of 4-MUG on the transport of E217βG. Transport of [3H]E217βG (50 nM for 30 s at 37°C) was examined in membrane vesicles expressing rat Mrp2 (○; A), dog Mrp2 (▪; B), and GFP (data not shown) in the presence of ATP. Each symbol represents the Mrp2-dependent transport, determined by subtracting the transport into GFP-expressing vesicles from that into respective Mrp2-expressing vesicles. Each point and vertical bar represents the mean ± S.D. of triplicate determinations. A broken line and a solid line represent fitting result of single site model and two-site model, respectively.
IC50 values of 4-MUG on the transport of [3H]E217ßG
Each value was calculated by fitting the data shown in Fig. 7. r represents the ratio of the contribution of the higher affinity component into the equations (see Materials and Methods). IC50High and IC50Low mean the IC50 value for the high-affinity site and for the low-affinity site, respectively. Values represent the obtained parameter ± computer calculated S.D.
Expression Levels of Rat and Dog Mrp2 Protein in the Liver. The Mrp2 protein expression level was compared in the crude membrane fraction from rat and dog liver in comparison with the standard MBP fusion proteins (Fig. 8). As shown in Fig. 8, the expression of Mrp2 protein in dog liver was quite low and calculated to be 10%, at most, of that in rat liver. The large difference in the Mrp2 protein expression was not attributable to any experimental artifacts because similar enrichment factors and also the absolute activity of the canalicular marker enzyme (alkaline phosphatase) were confirmed, as shown in Table 3.

The protein expression levels of Mrp2 in rat and dog liver. The homogenate and crude membrane prepared from rat and dog liver (10 μg of protein) were separated on an 8.5% polyacrylamide slab gel containing 0.1% SDS. The fractionated proteins were transferred to a membrane filter by electroblotting. The rat and dog Mrp2 were detected by anti-human MRP2 antiserum. The arrowhead represents Mrp2 band position (190 kDa).
The expression level of Mrp2 protein in rat and dog liver
Mrp2 protein expression levels in rat and dog livers were quantified by Western blot analysis using respective MBP fusion protein as a standard. Results represent the mean ± S.D. of triplicate determinations. The alkaline phosphatase (ALP) activities in the crude membrane fraction and liver homogenate are shown.
Discussion
There are two reported cDNA sequences of dog Mrp2 in the public database (Genbank accession No. Y18220 and AJ315401) (Conrad et al., 2001). The former (Y18220) has a shorter 4506-bp ORF sequence encoding a 1502 deduced amino acid sequence, and the latter (AJ315401) has a longer 4632-bp ORF sequence encoding a 1544 deduced amino acid sequence. These are considered splicing variants in the carboxy-terminal region. We were able to amplify dog Mrp2 with longer ORF (AJ315401) from beagle dog liver using a specific PCR primer set. We also tried to amplify the shorter variant using another specific primer set specifically designed for shorter variants, but no amplification took place (data not shown). This indicated that our longer ORF variant (encoding 1544 amino acids) is the major Mrp2 transcript in the liver of the beagle dog compared with the shorter one. This is consistent with the size of Mrp2 in other species: 1545 amino acids for human (Taniguchi et al., 1996), 1541 amino acids for rat (Büchler et al., 1996; Paulusma et al., 1996; Ito et al., 1997), 1543 amino acids for mouse (Fritz et al., 2000), 1544 amino acids for monkey, and 1564 amino acids for rabbit (van Kuijck et al., 1996). Actually, our dog Mrp2 cDNA encoded functional amino acid sequences as demonstrated in the vesicle transport study using glutathione and glucuronide conjugates as substrates (Fig. 4). Our dog Mrp2 cDNA contains 10 nucleotide substitutions that lead to seven amino acid substitutions compared with the reported sequence (protein accession no. CAC48162). Of these substitutions, an amino acid residue 1109 was in the predicted transmembrane region, and the others were in the predicted cytoplasmic region. Mutations or deletions in human MRP2 found in patients suffering from Dubin-Johnson syndrome affect its function or protein folding (Suzuki and Sugiyama, 2002). Moreover, the site-directed mutagenesis of some charged amino acids in the transmembrane region of human and rat Mrp2 affected their transport activity or substrate specificity (Ryu et al., 2000; Ito et al., 2001a,b,c). Since our seven amino acid substitutions did not correspond to these functionally relevant spots, such effects would be the minimum in our dog Mrp2. It should be elucidated whether the dog Mrp2 isolated in our study is the major genotype or not, as we used only one beagle dog liver for cloning material.
We investigated the transport characteristics of dog Mrp2 in comparison with rat Mrp2 using [3H]LTC4 as a glutathione conjugate and [3H]E217βG as a glucuronide conjugate (Fig. 4). Ishizuka et al. (1999) demonstrated that the transport activity of DNP-SG in rat CMVs was approximately 10-fold higher than that in dog CMVs (rat, 64.2 μl/min/mg protein; dog, 7.7 μl/min/mg protein), the difference in transport activities between rat and dog mainly depended on the Vmax values (Ishizuka et al., 1999). In our expression system, the ATP-dependent transport of [3H]LTC4 and [3H]E217βG normalized by the Mrp2 expression levels was similar in rat and dog. This observation suggested that the large functional difference previously observed in CMVs from rat and dog liver might be attributable to the expression level of Mrp2 protein rather than the intrinsic activity of the transporter molecule. Supporting this notion, the expression of Mrp2 in dog liver was less than 10% that in rat liver. Conrad et al. (2001) reported that the tissue distribution pattern of Mrp2 was similar to that in rat and human; namely, it was found in the liver, kidney, and small intestine (Conrad et al., 2001). However, in contrast to rat (Büchler et al., 1996; Paulusma et al., 1996; Ito et al., 1997; Paulusma and Oude Elferink, 1997), mouse (Fritz et al., 2000), and human (Taniguchi et al., 1996), the expression level in the liver was lower than in the kidney in dog (Conrad et al., 2001). Such lower Mrp2 expression in the liver was also reported in rabbit (small intestine>kidney≫liver) (van Kuijck et al., 1996). Moreover, in monkey, Mrp2 mRNA and protein are hardly detectable in the liver without treatment with tamoxifen or rifampin, both of which are inducers of Mrp2 (Kauffmann et al., 1998). The dog can be classified into a species whose Mrp2 expression in the liver is relatively low. These findings correspond well to the reported rank order of in vivo biliary excretion clearance of temocaprilat and the in vitro transport activity of DNP-SG into CMVs in rat, mouse, rabbit, and dog (Ishizuka et al., 1999). Quantitative analysis of the intrinsic transport activity of each Mrp2 from other species (mouse, rabbit, monkey, guinea pig, and human) will accomplish the precise mechanism underlying the large species difference in the biliary excretion potential of Mrp2 substrates. There is also a large species difference in the BSIDF (Erlinger, 1982). The rank order of the BSIDF was rat (70 μl/min/kg body weight), rhesus monkey (7 μl/min/kg body weight), dog (5 μl/min/kg body weight), and human (1.5–2 μl/min/kg body weight) (Erlinger, 1982), and this correlates well with the reported rank order of Mrp2 transport activity (Ishizuka et al., 1999). Moreover, the biliary excretion clearance of temocaprilat, a known Mrp2 substrate, was extremely higher in rat than in dog (rat; 39.2 ml/min/kg body weight versus dog; 0.8 ml/min/kg body weight) (Ishizuka et al., 1999). Our present study and a previous report (Ishizuka et al., 1999) highlight the importance of Mrp2 for the species differences in overall bile flow formation.
The ATP-dependent transport of LTC4 by dog Mrp2 was saturable with a Km value of 0.53 μM, which was comparable with that by rat Mrp2 [2.05 μM in our study and 1.1 μM in Cui et al. (1999)] and human MRP2 [1.0 μM in Cui et al. (1999)]. There is little species difference in the transport activity and affinity as far as LTC4 transport is concerned.
Although the initial transport rate does not seem to differ between rat and dog, a marked difference was observed when the concentration dependence experiment was performed for [3H]E217βG transport. The ATP-dependent transport of [3H]E217βG was only partially saturated in the presence of a maximum 75 μM E217βG. Since it was difficult to use a higher concentration of E217βG because of its solubility limit, data were fitted to a model with one saturable component (Km, Vmax) and nonsaturable component (PSns). In rat, a similar nonsaturable component was also observed (Fig. 6 and Table 1) but was so small and might be hardly detectable in the previous observations (Ito et al., 1998, 2001b; Cui et al., 1999). The low-affinity transport site for glucuronide compound was further characterized by the use of 4-MUG as a inhibitor for the transport of tracer concentration of [3H]E217βG (50 nM). 4-MUG is also a substrate of Mrp2, as demonstrated by using CMV from rats (Niinuma et al., 1997), and its solubility is high enough to cover the mM range. In the presence of 3 mM 4-MUG, ATP-dependent transport of [3H]E217βG was completely inhibited, indicating that the apparently nonsaturable component observed in the self-inhibition study (Fig. 6B) was really an Mrp2-dependent transport for both rat and dog (Fig. 7). Supporting this consideration, vesicle-associated [3H]E217βG transport was wholly osmosensitive in both rat and dog Mrp2. Actually, the two-site model was more appropriate to fit the data for dog Mrp2 with IC50High of 70.1 ± 51.8 μM and IC50Low of 1.54 ± 0.16 mM, respectively, and the contribution of these higher and lower affinity sites were 42% and 58%, respectively, for dog Mrp2 (Table 2). On the other hand, we could not see any advantage in fitting the data with the two-site model for rat Mrp2 (Fig. 7A). This may be due to less contribution of the lower affinity site for the glucuronide compound in rat Mrp2 (20% as estimated from Fig. 6B). Whereas the Km values were comparable between rat and dog, the V′max and PS′ns values normalized by the Mrp2 expression level were different. Consequently, the relative contribution of the high-affinity saturable site (V′max/Km) to the total transport (V′max/Km + PS′ns) gave a large difference at tracer concentrations of E217βG (80% and only 35% in rat and dog Mrp2, respectively). Multiple recognition sites on Mrp2 molecules have been proposed previously (Bakos et al., 2000; Evers et al., 2000; Ito et al., 2001b; Bodo et al., 2003; Zelcer et al., 2003); however, these consist of transport sites, and the others are modulation (inhibitory or stimulatory) site(s). As far as we know, our present observation is the first to implicate that Mrp2 has high- and low-affinity transport sites for glucuronide conjugates.
In conclusion, the large difference in the biliary excretion clearance of organic anions between rat and dog depends on the protein expression levels of Mrp2 in the liver. We found an additional low-affinity site that made a significant contribution to the transport of glucuronide conjugates in dog Mrp2. The unique transport properties of dog Mrp2 possibly cause a species difference, especially in Mrp2-based drug-drug interactions or drug-induced hyperbilirubinemia.
Footnotes
-
This work was supported in part by a Grant-in-Aid for Scientific Research (15790085) from Japan Society for the Promotion of Science.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.104.002196.
-
ABBREVIATIONS: Mrp, multidrug resistance-associated protein; BSIDF, bile salt-independent flow; bp, base pair(s); CMVs, canalicular membrane vesicles; DNP-SG, 2,4-dinitrophenyl-S-glutathione; E217βG, 17β-estradiol 17-(β-D-glucuronide); LTC4, leukotriene C4; 4-MUG, 4-methylumbelliferyl-β-D-glucuronide; PCR, polymerase chain reaction; GFP, green fluorescent protein; BSA, bovine serum albumin; PSns, nonsaturable component; AIC, Akaike’s information criterion; ORF, open reading frame; MBP, maltose binding protein.
- Received September 4, 2004.
- Accepted October 22, 2004.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}