


Abstract
Flurbiprofen is a nonsteroidal anti-inflammatory drug used as a racemic mixture. Although glucuronidation is one of its elimination pathways, the role of UDP-glucuronosyltransferase (UGT) in this process remains to be investigated. Thus, the kinetics of the stereoselective glucuronidation of racemic (R,S)-flurbiprofen by recombinant UGT isozymes and human liver microsomes (HLMs) were investigated, and the major human UGT isozymes involved were identified. UGT1A1, 1A3, 1A9, 2B4, and 2B7 showed glucuronidation activity for both (R)- and (S)-glucuronide, with UGT2B7 possessing the highest activity. UGT2B7 formed the (R)-glucuronide at a rate 2.8-fold higher than that for (S)-glucuronide, whereas the other UGTs had similar formation rates. The glucuronidation of racemic flurbiprofen by HLMs also resulted in the formation of (R)-glucuronide as the dominant form, which occurred to a degree similar to that by recombinant UGT2B7 (2.1 versus 2.8). The formation of (R)-glucuronide correlated significantly with morphine 3-OH glucuronidation (r = 0.96, p < 0.0001), morphine 6-OH glucuronidation (r = 0.91, p < 0.0001), and 3′-azido-3′-deoxythymidine glucuronidation (r = 0.85, p < 0.0001), a reaction catalyzed mainly by UGT2B7, in individual HLMs. In addition, the formation of both glucuronides correlated significantly (r = 0.99, p < 0.0001). Mefenamic acid inhibited the formation of both (R)- and (S)-glucuronide in HLMs with similar IC50 values (2.0 and 1.7 μM, respectively), which are close to those in recombinant UGT2B7. In conclusion, these findings suggest that the formation of (R)- and (S)-glucuronide from racemic flurbiprofen is catalyzed by the same UGT isozyme, namely UGT2B7.
Flurbiprofen [(±)-(R,S)-2-(2-fluoro-4-biphenyl)propionic acid] (Fig. 1) is commonly used as an analgesic nonsteroidal anti-inflammatory drug. Although the anti-inflammatory activity is almost entirely attributable to (S)-flurbiprofen (Kulmacz and Lands, 1985), it has been suggested that both (R)- and (S)-flurbiprofen may possess antinociceptive activity (Brune et al., 1991; Geisslinger and Schaible, 1996). In addition, both enantiomers have been shown to exhibit antiproliferative effects (McCracken et al., 1996). Flurbiprofen exists as a chiral compound with a stereoselective disposition in humans (Geisslinger et al., 1994) and is metabolized via several drug metabolizing pathways, including cytochromes P450 and UDP-glucuronosyltransferases (UGTs). Approximately 60 to 70% of the administered dose is eliminated as acyl glucuronides (Davies, 1995). After oral administration of racemic flurbiprofen (100 mg/kg), 8.4 and 7.3% of the dose was excreted into the urine as the acyl glucuronide of (R)- and (S)-flurbiprofen, respectively (Patel et al., 2003). However, the flurbiprofen glucuronidation fraction is difficult to determine and may be higher because of the instability of acyl glucuronides in vivo. The data from these reports suggest that the formation of glucuronide constitutes some portion of the metabolic pathway of flurbiprofen. The other major oxidative metabolite (4′-hydroxyflurbiprofen) and minor oxidative metabolite (3′-hydroxy-4′-methoxyflurbiprofen) were also detected in urine. Although it is known that cytochrome P450 2C9 is the main enzyme involved in the cytochrome P450-mediated oxidation of flurbiprofen (Tracy et al., 1995), the UGTs responsible for flurbiprofen glucuronidation remain to be determined. Previous findings demonstrated that (S)-flurbiprofen was an effective UGT2B7 inhibitor (Bauman et al., 2005), and the recombinant human UGTs, 1A1, 1A3, 1A4, 1A7, 1A8, 1A9, 1A10, 2B4, 2B7, and 2B15, have all been shown to catalyze racemic flurbiprofen glucuronidation (Kuehl et al., 2005). However, no enantioselective assay for flurbiprofen glucuronidation has been conducted as of yet, and the main UGT isozymes responsible for the formation of (R)- and (S)-flurbiprofen glucuronide remain to be determined. The novel information presented in the current study includes the identification of the predominant UGT isozymes involved in flurbiprofen glucuronidation in the human liver, and the determination of whether the UGT isozymes involved in the glucuronidation of the (R) and (S) forms differs. A simple comparison of the glucuronidation activity among UGT isozymes is not sufficient to elucidate the main UGTs involved in the liver because the UGT content in each recombinant UGT isozyme could not be determined, and the relative abundance of UGTs in the human liver has not been determined. In addition, because little data on the specific inhibitors of the respective UGT isozyme are available, identification of the main UGT isozymes involved in the glucuronidation of drugs is more difficult than that in cytochrome P450-mediated metabolism. Thus, to determine the responsible UGT isozyme, correlation studies as well as inhibition studies should be conducted. The glucuronidation of β-estradiol at the 3-OH position and propofol were used as probe reactions for UGT1A1 and 1A9, respectively (Mano et al., 2006, 2007a), whereas the glucuronidation of morphine and 3′-azido-3′-deoxythymidine (AZT) were used as typical UGT2B7 reactions (Court et al., 2003; Mano et al., 2007b). Thus, correlation studies between these probe reactions and the glucuronidation of the drugs of interest in liver microsomes from individual humans, together with inhibition characteristics, should help in the determination of the UGT isozymes responsible. Here, we investigated the main UGT isozymes involved in flurbiprofen glucuronidation in the human liver using human liver microsomes (HLMs) and recombinant human UGT isozymes.

Chemical structures of (R)- and (S)-flurbiprofen.
Materials and Methods
Chemicals and Reagents. Racemic flurbiprofen was purchased from Sigma (St. Louis, MO). A panel of recombinant human UGT Supersomes (UGT1A1, 1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2B4, 2B7, 2B15, and 2B17) expressed in baculovirus-infected insect cells were purchased from BD Biosciences (Bedford, MA). Pooled and individual HLMs were purchased from Xenotech LLC (Kansas City, KS). Because this study uses materials from humans, approval was obtained from the Astellas Pharma Inc. Ethics Committee. All other chemicals were of the highest grade.
Kinetic Study in HLMs. Racemic flurbiprofen was incubated for 20 min in 0.25 ml of a reaction mixture consisting of 50 mM Tris-HCl buffer (pH 7.5) containing 8 mM MgCl2, 25 μg/ml alamethicin, 10 mM saccharic acid 1,4-lactone, and HLMs in the absence or presence of 5 mM UDP-glucuronic acid (UDPGA). Saccharic acid 1,4-lactone (10 mM) increased the formation of both (R)- and (S)-glucuronide 2.0- and 1.6-fold, respectively. The substrate concentration was set at 1 to 400 μM, and the protein concentration was 0.2 mg of protein/ml. After preincubation of the reaction mixtures for 5 min at 37°C, the reaction was started by adding UDPGA (5 mM), followed by incubation for 20 min at 37°C. The kinetic studies were conducted using protein concentrations and incubation times that yielded linear product formation. The reactions were terminated by adding acetonitrile (0.05 ml), 0.01 ml of 10% formic acid (v/v), and 0.02 ml of internal standard (sulindac, 50 μM), followed by centrifugation at 1870g for 10 min to obtain the supernatant. Aliquots (0.12 ml) of the supernatant were injected into a high-performance liquid chromatography (HPLC) equipped with an ultraviolet (UV) detector.
Panel Study by UGT Isozymes. The formation of both (R)- and (S)-glucuronide from racemic flurbiprofen was measured in recombinant human UGTs 1A1, 1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2B4, 2B7, 2B15, and 2B17. The concentrations of the substrate and the UGT isozymes were 50 μM and 0.5 mg of protein/ml, respectively. Reaction time was 60 min for all isozymes.
Assay. The peak areas of (R)- and (S)-flurbiprofen glucuronide were analyzed using reverse-phase HPLC according to the reported method (Mano et al., 2002). In brief, the HPLC system consisted of LC-10AS pumps, a SCL-10A system controller, a SIL-10A autosampler, a SPD-10AV UV detector, and a C-R4AX integrator (Shimadzu, Kyoto, Japan). Chromatographic separation was achieved using a Capcellpak UG80 column (4.6 × 150 mm, 5 μm; Shiseido, Tokyo, Japan). The mobile phase consisted of 20 mM ammonium acetate (pH 5.6)/ethanol/acetonitrile [20:7:2 (v/v)] and was delivered at a flow rate of 1.0 ml/min. The retention times of (R)- and (S)-glucuronide, detected at a UV wavelength of 240 nm, were approximately 15 and 16 min, respectively, which were identified by reference to the previous report (Mano et al., 2002). Because authentic standards for (R)- and (S)-glucuronide were not available, unknown concentrations of the glucuronides were determined by reference to a standard curve for racemic flurbiprofen. The standard curve for flurbiprofen was linear over the concentration range of 0.5 to 10 μM, with correlation coefficient values of >0.99. The limit of quantification for the glucuronidation assay was 19 pmol equivalents/min/mg protein, and the precision and accuracy were less than 15%.
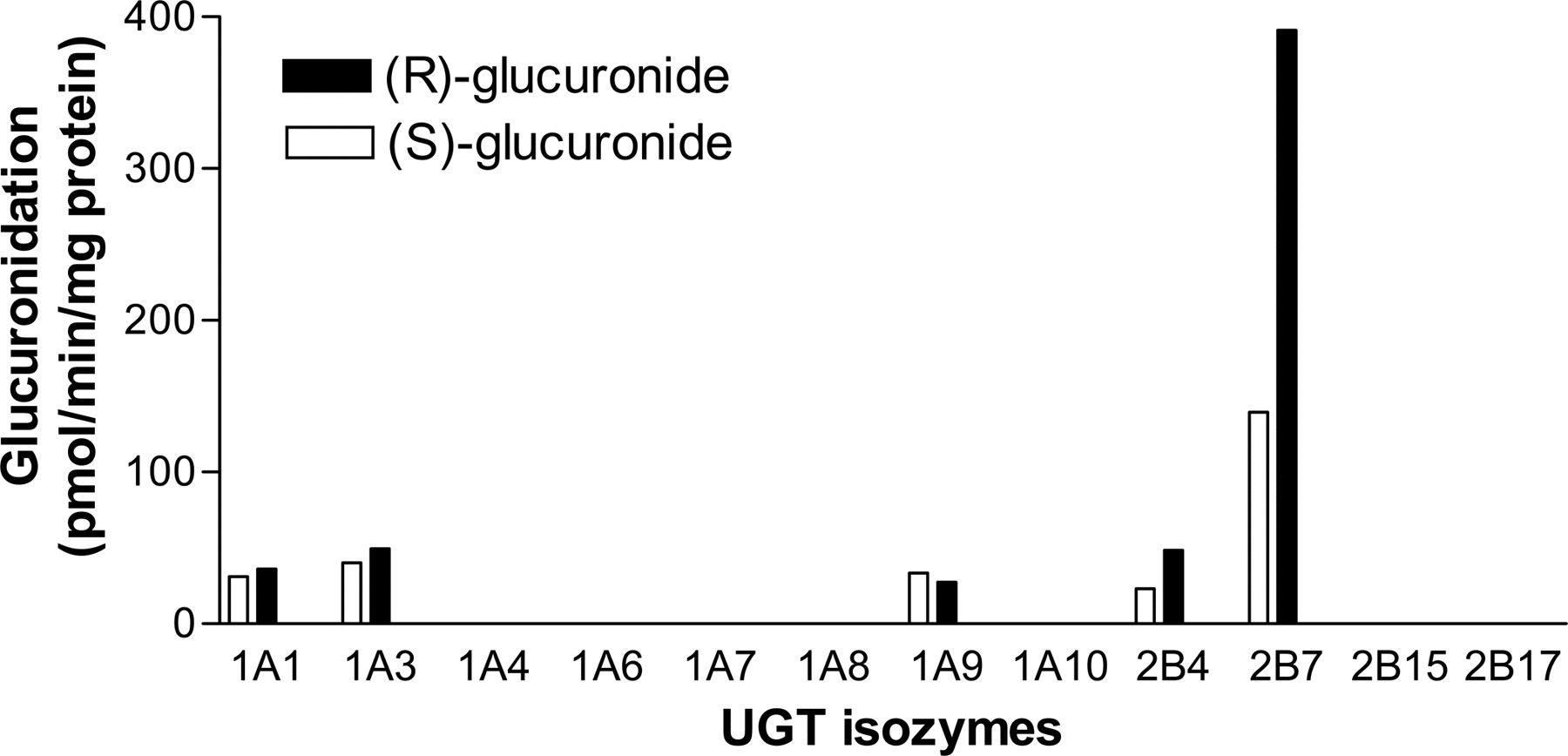
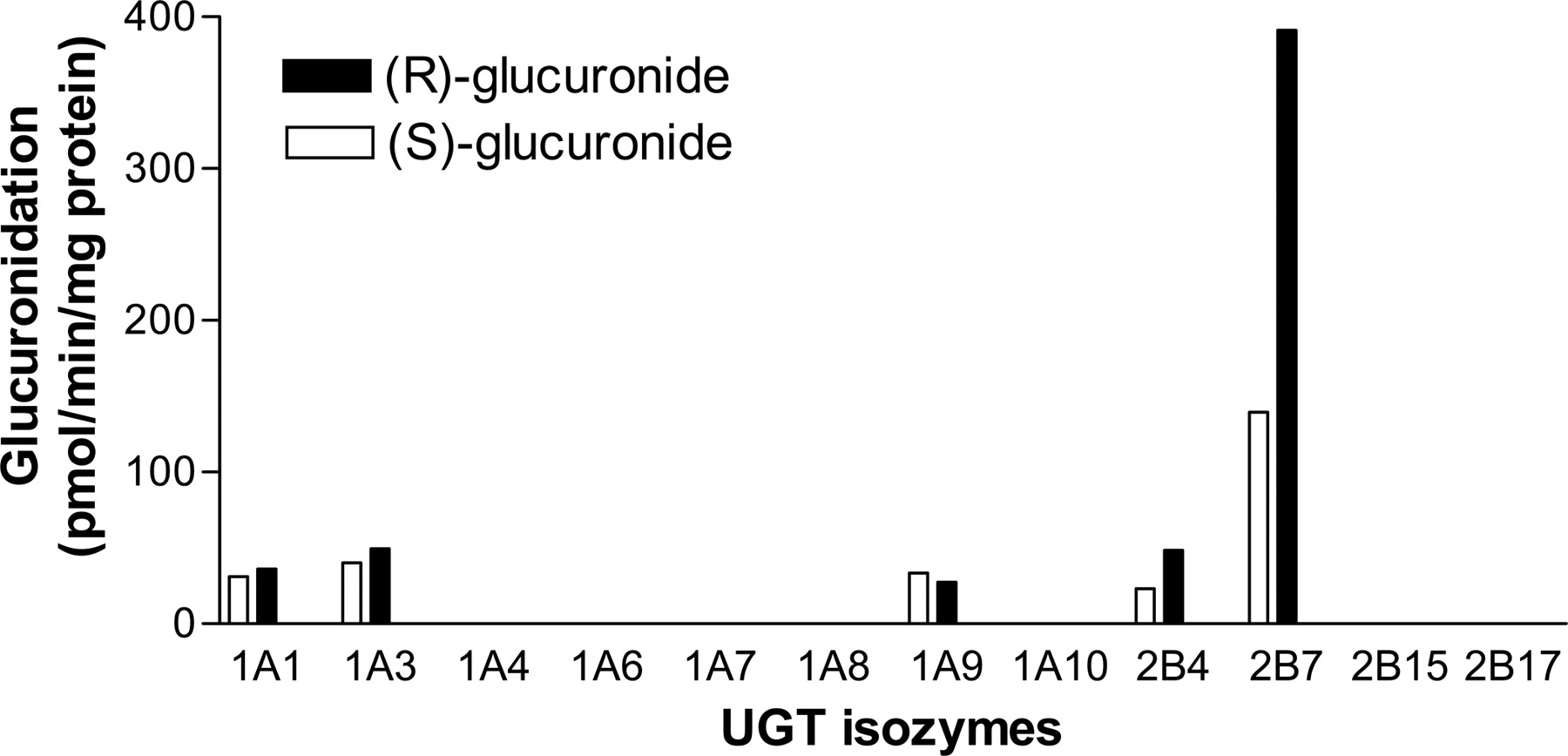
Formation of (R)- and (S)-flurbiprofen glucuronides from racemic flurbiprofen by recombinant human UGT isozymes. Racemic flurbiprofen (50 μM) was incubated with recombinant human UGT isozymes (UGT1A1, 1A3, 1A4, 1A6, 1A7, 1A8, 1A9, 1A10, 2B4, 2B7, 2B15, and 2B17) at the protein concentration of 0.5 mg of protein/ml for 60 min. Open and closed columns represent the (S)- and (R)-glucuronides formation, respectively, and the data represent the mean of duplicate determinations. The limit of quantification for the flurbiprofen glucuronidation assay was 19 pmol equivalents/min/mg protein.
Correlation Study. Flurbiprofen glucuronidation was measured in microsomes from 16 individual human livers. Racemic flurbiprofen (20 μM) was incubated with HLMs (0.2 mg of protein/ml) for 20 min at 37°C. The correlation between the formation of (R)- and (S)-glucuronide and the glucuronidation of β-estradiol at the 3-OH position, propofol, morphine at the 3- and 6-OH positions, and AZT was determined using Pearson's moment method and Prism version 3.02 (GraphPad Software, San Diego, CA). The manufacturer used the glucuronidation activity of β-estradiol at the 3-OH position, propofol, and morphine at the 3- and 6-OH positions in the microsomes from 16 human livers as typical reference activities for UGT1A1, 1A9, and 2B7, respectively. AZT glucuronidation was measured by a previously reported method (Mano et al., 2007b).
Chemical Inhibition. Mefenamic acid, a potent inhibitor of UGT2B7-catalyzed AZT glucuronidation (Mano et al., 2007b), was tested for its inhibitory effect on flurbiprofen glucuronidation in HLMs and recombinant UGT2B7. Racemic flurbiprofen (20 or 50 μM) was incubated with HLMs and recombinant UGT2B7 (0.2 mg of protein/ml) in the presence or absence of mefenamic acid (0.1–10 μM) to determine the IC50 value of mefenamic acid. No interference by the inhibitor or its glucuronide was detected in the assay of the flurbiprofen glucuronides.
Data Analysis. The kinetics of glucuronidation by HLMs were fitted to Michaelis-Menten kinetics with a nonsaturable component (eq. 1) to estimate the Michaelis constant (Km), maximum velocity (Vmax), and nonsaturable clearance (CLns) (Mano et al., 2006). These calculations were carried out using Prism version 3.02. The intrinsic clearance (CLint) value was calculated from Vmax/Km + CLns.  The IC50 value, which represents the concentration of the inhibitor that will inhibit 50% of the activity of the control, was estimated using Prism version 3.02.
The IC50 value, which represents the concentration of the inhibitor that will inhibit 50% of the activity of the control, was estimated using Prism version 3.02.
Results
Glucuronidation by UGT Isozymes. UGT1A1, 1A3, 1A9, 2B4, and 2B7 all catalyzed the glucuronidation of both (R)- and (S)-flurbiprofen, with UGT2B7 having the highest activity (Fig. 2). For UGT1A1, 1A3, 1A9, 2B4, and 2B7, the (R)-glucuronidation velocity values were 36, 49, 27, 48, and 391 pmol equivalents/min/mg protein, respectively, and the (S)-glucuronidation values were 31, 40, 34, 23, and 139 pmol equivalents/min/mg protein, respectively. All other recombinant UGT isozymes had activity values of less than 19 pmol equivalents/min/mg protein.

Kinetics for the formation of (R)- and (S)-flurbiprofen glucuronide by incubation of racemic flurbiprofen with HLMs. Racemic flurbiprofen (1–400 μM) was incubated with protein concentrations of 0.2 mg of protein/ml for 20 min in HLMs. Data represent the both duplicate data points. The Eadie-Hofstee plot is presented in the inset.
Glucuronidation by HLMs. The incubation of flurbiprofen with HLMs resulted in two peaks, which were observed only in the presence of UDPGA. The formation of (R)- and (S)-flurbiprofen glucuronide from racemic flurbiprofen yielded Michaelis-Menten kinetics with a nonsaturable component and Km values of 12 ± 1.1 and 7.5 ± 1.1 μM, respectively (Fig. 3, mean ± computer-calculated S.E.). The respective Vmax values were 1483 ± 49 and 454 ± 19 pmol equivalents/min/mg protein, and the respective CLns values were 5.8 ± 0.2 and 1.5 ± 0.1 μl/min/mg protein. The CLint values were 129 and 62 μl/min/mg protein for the formation of (R)- and (S)-glucuronide, respectively.
Correlation Studies in HLMs. The formation of (R)- and (S)-flurbiprofen glucuronide in microsomes from 16 human livers ranged from 750 to 2024 and 218 to 759 pmol equivalents/min/mg protein, respectively. As shown in Fig. 4, A–C, (R)-glucuronide formation correlated significantly with AZT glucuronidation (r = 0.85, p < 0.0001), the glucuronidation of morphine at the 3-OH position (r = 0.96, p < 0.0001), and the glucuronidation of morphine at the 6-OH position (r = 0.91, p < 0.0001), which are typical reactions for UGT2B7. In contrast, no significant correlation was observed with estradiol 3β-glucuronidation or propofol glucuronidation, which are probe reactions for UGT1A1 and 1A9, respectively (Fig. 4, D and E). The correlation between the formation of (R)-glucuronide and (S)-glucuronide was significant (Fig. 5, r = 0.99, p < 0.0001).
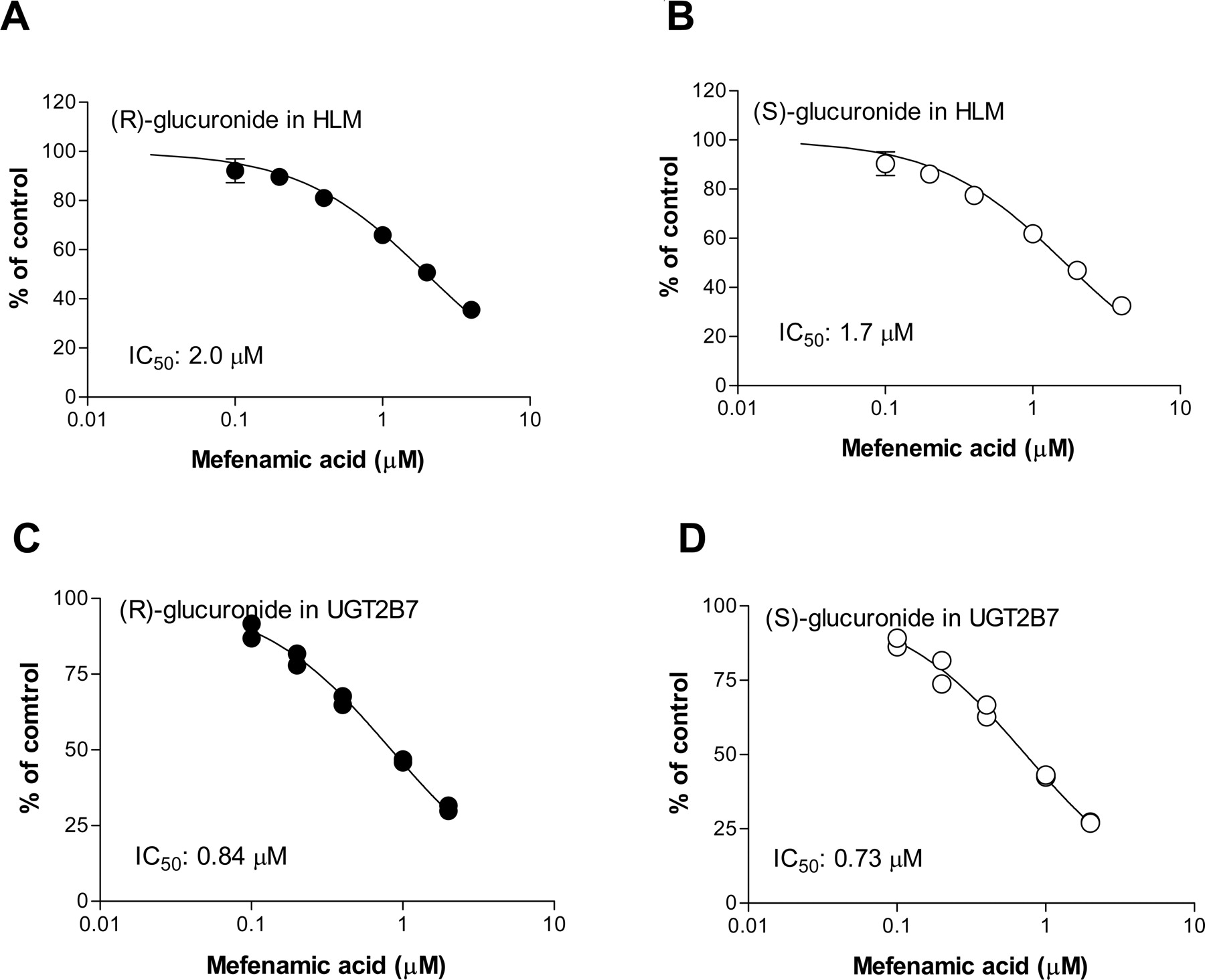
Chemical Inhibition. The inhibitory effect of mefenamic acid on flurbiprofen glucuronidation in HLMs was evaluated. As shown in Fig. 6, A and B, mefenamic acid inhibited both (R)- and (S)-glucuronide formation in HLMs in a concentration-dependent manner, with IC50 values of 2.0 ± 0.1 and 1.7 ± 0.1 μM (mean ± computer-calculated S.E.), respectively. The inhibitory effects of mefenamic acid on flurbiprofen glucuronidation were also evaluated in recombinant UGT2B7. The IC50 values of mefenamic acid in recombinant UGT2B7 were 0.84 μM for (R)-glucuronide and 0.73 μM for (S)-glucuronide (Fig. 6, C and D).
Discussion
The main UGT isozymes involved in flurbiprofen glucuronidation in the human liver were investigated in this study. The recombinant human UGT isozymes 1A1, 1A3, 1A9, 2B4, and 2B7 showed glucuronidation activity (Fig. 2). This finding is partially consistent with a previous report (Kuehl et al., 2005), in which UGT1A4, 1A6, 1A8, 1A10, and 2B17 also had slight glucuronidation activity. The reason for the discrepancy between the present and the previous findings may be due to differences in the detection sensitivities of HPLC-UV and liquid chromatography with tandem mass spectrometry. The ratio of (R)-glucuronide to (S)-glucuronide varied among the UGT isozymes, but (R)-glucuronide was the predominant form in recombinant UGT2B7 (formation ratio: 2.8), although the ratios in all UGT isozymes other than UGT2B7 were comparable (Fig. 2). The finding in this study is considered to be important because it strongly suggests that UGT2B7 is the primary UGT isozyme involved in flurbiprofen glucuronidation. Although the study by Kuehl et al. (2005) demonstrated that UGT2B7 showed the highest flurbiprofen glucuronidation activity among the UGT isozymes studied, these data did not lead to the conclusion that UGT2B7 is the main UGT isozyme responsible. This is because the amount of UGT in each recombinant UGT system and the relative abundance of UGT isozymes in the liver remain to be determined. In addition, this study represents the predominant glucuronidation of (R)-flurbiprofen, which is consistent with the results in humans given racemic flurbiprofen (Patel et al., 2003). Although the ratio of (R)-glucuronide to (S)-glucuronide varied among the UGT isozymes investigated, UGT1A1, 1A3, 1A9, 2B4, and 2B7 showed glucuronidation activity for both (R)- and (S)-flurbiprofen. This is consistent with the glucuronidation of (R,S)-propranolol, both forms of which were glucuronidated by UGT1A1, 1A9, 1A10, 2B4, 2B7, and 2B17 (Sten et al., 2006). In contrast, stereoselective glucuronidation has been reported for (R,S)-etodolac (Tougou et al., 2004) and (R,S)-oxazepam (Court et al., 2002). UGT1A7, 1A8, 1A10, and 2B15 showed glucuronidation activity for (S)-etodolac but not (R)-etodolac (Tougou et al., 2004). UGT1A1, 1A6, and 2B15 showed glucuronidation activity only for (S)-oxazepam, whereas UGT1A7 and 1A9 catalyzed only (R)-oxazepam glucuronidation (Court et al., 2002).
UGT1A1, 1A3, 1A9, 2B4, and 2B7 all exhibit glucuronidation activity and are expressed in the human liver, which suggests that they may be involved with flurbiprofen glucuronidation in the liver. As flurbiprofen is used as a racemate in the clinical setting, the kinetic study for the glucuronidation of racemic flurbiprofen was conducted in HLMs. The Eadie-Hofstee plot shown in Fig. 3 suggests that there is more than one isozyme involved in flurbiprofen glucuronidation. The CLint value for (R)- and (S)-glucuronide formation was 129 and 62 μl/min/mg protein, and the respective CLns values were 5.8 and 1.5 μl/min/mg protein. This finding indicates that the high affinity component contributes significantly to the total CLint value. As was observed in recombinant UGT2B7, (R)-glucuronide was also the predominant form in HLMs. The ratio of CLint for the formation of (R)-glucuronide to (S)-glucuronide is 2.1 in HLMs, which is similar to that in recombinant UGT2B7 (2.8). The plasma levels of (R)-flurbiprofen were lower than those of (S)-flurbiprofen in humans given racemic flurbiprofen with the plasma area under the curve ratio of (R)-flurbiprofen to (S)-flurbiprofen ranging from 0.79 to 0.88 (Davies, 1995). In addition, the plasma levels of (R)-flurbiprofen glucuronide were higher than those of (S)-flurbiprofen glucuronide, with a S/R ratio of 0.74 (Patel et al., 2003). This ratio is somewhat higher than that in vitro [in HLMs (0.48)], but the in vivo plasma levels of (R)- and (S)-flurbiprofen glucuronide are influenced by other processes such as plasma protein binding and excretion clearance. The kinetic profiles in HLMs obeyed Michaelis-Menten kinetics with Km values of 12 and 7.5 μM for the formation of (R)- and (S)-glucuronide, respectively (Fig. 3). The Km value in this study was somewhat lower than that in the previous study (56.3 μM; Kuehl et al., 2005). This discrepancy may be due to differences in the reaction conditions, because the substrate concentration differed between the former and present studies (10–1000 and 1–400 μM, respectively). In addition, the kinetic equations used to estimate the kinetic parameters were also different; no nonsaturable component was included in the equation used in the former study.
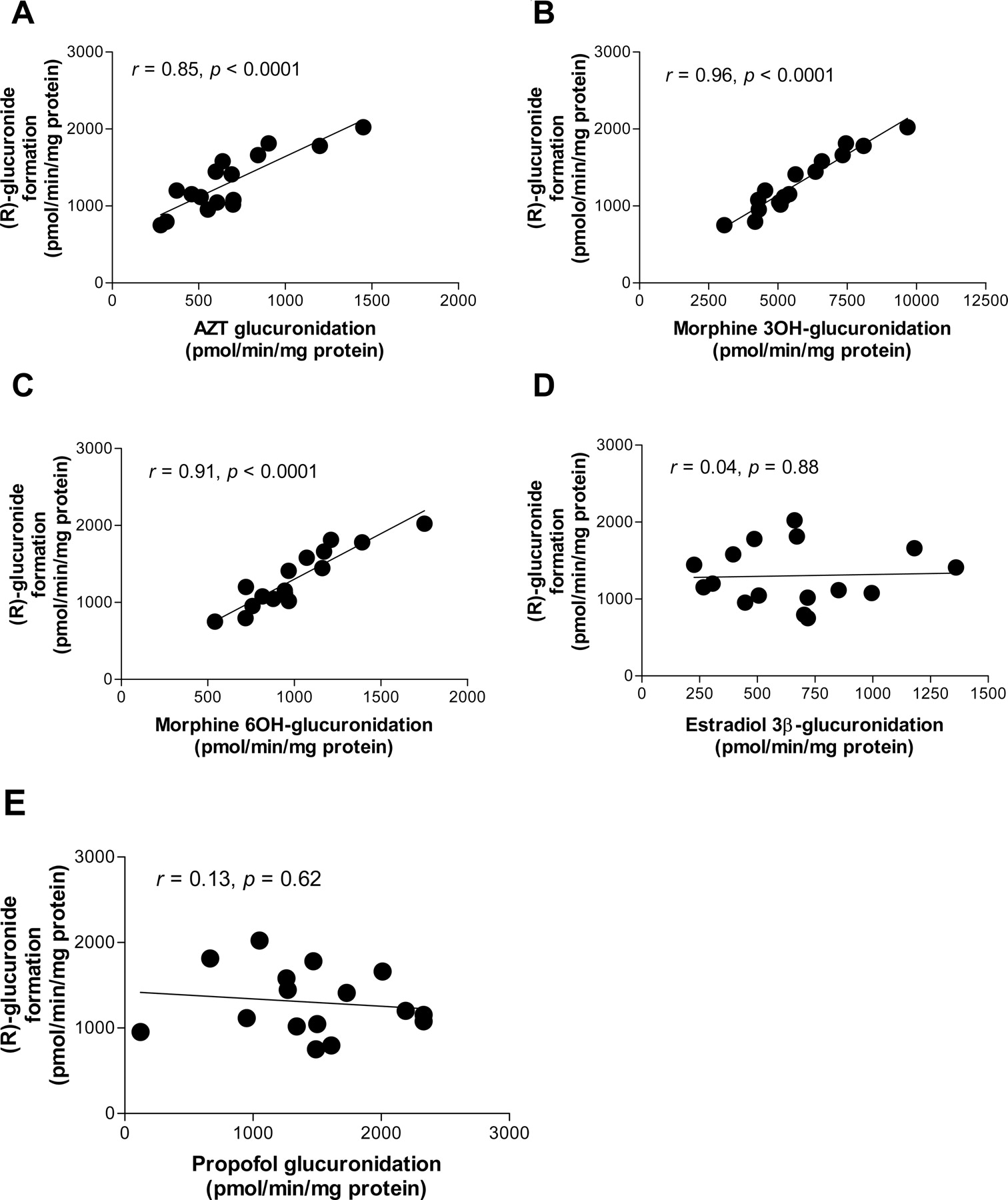
Correlation analysis between flurbiprofen glucuronidation and UDP-glucuronosyltransferase isozyme-catalyzed glucuronidation in microsomes from 16 individual human livers. Racemic flurbiprofen (20 μM) was incubated with microsomes (0.2 mg of protein/ml) for 20 min. Data represent the mean of duplicate determinations. The x-axis represents the activity for AZT glucuronidation (A), morphine 3-OH glucuronidation (B), morphine 6-OH glucuronidation (C), estradiol 3β-glucuronidation (D), and propofol glucuronidation (E); the y-axis represents the formation velocity of the (R)-glucuronide of flurbiprofen.

Correlation between the formation of (R)- and (S)-glucuronides in microsomes from 16 individual human livers. Racemic flurbiprofen (20 μM) was incubated with microsomes (0.2 mg of protein/ml) for 20 min. Data represent the mean of duplicate determinations. The x-axis represents the formation of (R)-glucuronide, and the y-axis represents that of (S)-glucuronide.
Because the ratio of (R)-glucuronide to (S)-glucuronide in UGT2B7 was found to be comparable with that in HLMs (2.8 versus 2.1), a correlation analysis between the glucuronidation activity of flurbiprofen and those of UGT2B7-specific morphine (3-OH and 6-OH) and AZT (Court et al., 2003; Mano et al., 2007b) was performed using microsomes prepared from 16 individual human livers to assess the significance of the contribution UGT2B7 makes to flurbiprofen glucuronidation. The formation of both (R)- and (S)-glucuronide correlated significantly with morphine 3-OH glucuronidation and 6-OH glucuronidation and AZT glucuronidation (Fig. 4, A–C); however, no significant correlation was observed for estradiol 3β-glucuronidation or propofol glucuronidation (Fig. 4, D and E), which are typical reactions for UGT1A1 and 1A9, respectively. These findings suggest that UGT2B7 is the predominant UGT isozyme responsible for flurbiprofen glucuronidation in the human liver. Although several UGT isozymes other than UGT2B7 are involved in morphine 3-OH and 6-OH glucuronidation (Court et al., 2003), the correlation coefficient for the UGT2B7 protein content between morphine 3-OH and 6-OH glucuronidation is statistically significant, and the values are comparable (0.50 and 0.51, respectively). Thus, as Court et al. (2003) suggest, either glucuronidation type can serve as a probe reaction for UGT2B7 as well as AZT glucuronidation. Figure 4 shows that the correlation coefficient of AZT and morphine 3-OH and 6-OH glucuronidation is highly significant and similar (0.85, 0.96, and 0.91, respectively). A correlation study in HLMs with a probe substrate for UGT1A3 is useful for eliminating this isozyme as an important contributor to flurbiprofen glucuronidation. However, the probe substrate for UGT1A3 is not fully understood, and only F6-1α, 23S, 25-trihydroxyvitamin D3 has been reported as a possible probe substrate for this isozyme (Kasai et al., 2005), which is not a commercially available substrate. For this reason, a correlation study using a probe reaction for UGT1A3 could not be conducted. Because it has been reported that the mRNA level of UGT1A3 was approximately 20-fold less than that of UGT1A1 (Mojarrabi et al., 1996), it is unlikely that UGT1A3 would play a significant role in flurbiprofen glucuronidation.

Inhibitory effects of mefenamic acid on flurbiprofen glucuronidation in HLMs and recombinant UGT2B7. Racemic flurbiprofen (20 or 50 μM) was incubated with HLMs and recombinant UGT2B7 in the absence and presence of mefenamic acid (0.1–10 μM), and the effects of mefenamic acid on the formation of (R)-glucuronide (A and C) and (S)-glucuronide (B and D) were assessed. The protein concentration was 0.2 mg of protein/ml, and the incubation time was 20 min. Data represent the mean ± S.D. of triplicate determinations in HLMs and both duplicate data points in recombinant UGT2B7.
The high correlation and low intercept for (R)- and (S)-glucuronide formation in 16 individual liver microsomes (Fig. 5) strongly suggests that these two glucuronides are formed by the same UGT isozyme. In addition, the inhibition study showed that mefenamic acid, a potent UGT2B7 inhibitor (Mano et al., 2007b), inhibited the formation of both (R)- and (S)-glucuronide with IC50 values of 2.0 and 1.7 μM, respectively, which are similar and close to that for recombinant UGT2B7 (Fig. 6, A–D). These findings led to the conclusion that UGT2B7 plays a predominant role in the formation of both (R)- and (S)-glucuronide of flurbiprofen in the human liver. Determination of the Km values for flurbiprofen glucuronidation in recombinant UGT1A3 and 2B4 have been helpful for confirming the minor involvement of these isozymes in flurbiprofen glucuronidation. However, differences in the glucuronidation Km values between recombinant UGT isozymes and HLMs are frequently observed, possibly due to heterodimerization of the UGT isozymes (Fujiwara et al., 2007). In addition, the glucuronidation activity for these UGT isozymes was low; thus, glucuronidation at the lower substrate concentrations could not be determined, which does not allow for a precise determination of kinetic parameters. This precluded any study of flurbiprofen glucuronidation kinetics in recombinant UGT isozymes.
Because UGT1A1, 1A3, 1A9, and 2B7, which exhibit flurbiprofen glucuronidation activity, are expressed in the intestine as well as the liver, intestinal glucuronidation should also be taken into consideration for full assessment of flurbiprofen glucuronidation. In conclusion, these findings indicate that UGT2B7 is the predominant UGT isozyme responsible for the glucuronidation of both enantiomers in the human liver.
Footnotes
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.107.015347.
-
ABBREVIATIONS: UGT, UDP-glucuronosyltransferase; AZT, 3′-azido-3′-deoxythymidine; HLM, human liver microsome; UDPGA, UDP-glucuronic acid; HPLC, high-performance liquid chromatography.
- Received February 23, 2007.
- Accepted April 16, 2007.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}