


Abstract
The aim of this study was to investigate the effect of commonly used botanicals on UDP-glucuronosyltransferase (UGT) 1A4, UGT1A6, and UGT1A9 activities in human liver microsomes. The extracts screened were black cohosh, cranberry, echinacea, garlic, ginkgo, ginseng, milk thistle, saw palmetto, and valerian in addition to the green tea catechin epigallocatechin gallate (EGCG). Formation of trifluoperazine glucuronide, serotonin glucuronide, and mycophenolic acid phenolic glucuronide was used as an index reaction for UGT1A4, UGT1A6, and UGT1A9 activities, respectively, in human liver microsomes. Inhibition potency was expressed as the concentration of the inhibitor at 50% activity (IC50) and the volume in which the dose could be diluted to generate an IC50-equivalent concentration [volume/dose index (VDI)]. Potential inhibitors were EGCG for UGT1A4, milk thistle for both UGT1A6 and UGT1A9, saw palmetto for UGT1A6, and cranberry for UGT1A9. EGCG inhibited UGT1A4 with an IC50 value of (mean ± S.E.) 33.8 ± 3.1 μg/ml. Milk thistle inhibited both UGT1A6 and UGT1A9 with IC50 values of 59.5 ± 3.6 and 33.6 ± 3.1 μg/ml, respectively. Saw palmetto and cranberry weakly inhibited UGT1A6 and UGT1A9, respectively, with IC50 values >100 μg/ml. For each inhibition, VDI was calculated to determine the potential of achieving IC50-equivalent concentrations in vivo. VDI values for inhibitors indicate a potential for inhibition of first-pass glucuronidation of UGT1A4, UGT1A6, and UGT1A9 substrates. These results highlight the possibility of herb-drug interactions through modulation of UGT enzyme activities. Further clinical studies are warranted to investigate the in vivo extent of the observed interactions.
Introduction
Conjugation of compounds with glucuronic acid represents a major disposition pathway for endogenous and exogenous compounds, including drugs and phytochemicals. Human glucuronidation enzymes [UDP-glucuronosyltransferases (UGTs)] are divided into two families, UGT1 and UGT2, which encompass more than 18 enzymes (Tukey and Strassburg, 2000). UGT1A4, UGT1A6, and UGT1A9 enzymes belong to the UGT1 family and conjugate a wide spectrum of drugs and phytochemicals. UGT enzymes are differentially expressed in tissues, with liver and intestine being the main sites for drug glucuronidation (Tukey and Strassburg, 2000). Substrates for UGT enzymes include many drugs (e.g., mycophenolic acid, trifluoperazine, tamoxifen, lamotrigine, and acetaminophen) and phytochemicals (e.g., quercetin, kaempferol, and epigallocatechin gallate) (Oliveira and Watson, 2000; Lu et al., 2003; Kiang et al., 2005). Because many phytochemicals are glucuronidated primarily by UGT1A enzymes, there is a potential for herb-drug interaction through competition with drug substrates for this pathway (Mohamed and Frye, 2011). We previously reported the inhibitory effects of several commonly used herbal supplements on UGT1A1 and of Ginkgo biloba extract and its polyphenolic compounds quercetin and kaempferol on UGT1A9 (Mohamed and Frye, 2010; Mohamed et al., 2010). The aim of this study was to identify other potential herb-UGT interactions through screening of commonly used herbal extracts for inhibitory effects on the activities of UGT1A4, UGT1A6, and UGT1A9.
Recent surveys estimate that 38% of Americans use complementary and alternative medicine, which includes herbal supplements (Barnes et al., 2008). However, the physiologic and metabolic effects of herbals and phytochemicals are often poorly understood. One of the issues of concern to clinicians is the potential for herb-drug interactions, which may lead to poor clinical outcomes (Gardiner et al., 2008). Several case studies have described deleterious herb-drug interactions that can lead to morbidity or even mortality (Ruschitzka et al., 2000; Kupiec and Raj, 2005). Therefore, much attention has been given to investigating the effects of herbal supplements on cytochrome P450 enzymes, the primary metabolic route for the majority of marketed drugs (Izzo and Ernst, 2009). In contrast, research regarding the potential of herbal products to alter other metabolic routes including glucuronidation is lacking (Mohamed and Frye, 2011).
Identification of selective substrates for UGT enzymes allows screening of herb-UGT interactions using human liver microsomes. Trifluoperazine, serotonin, and mycophenolic acid were reported to be selective in vitro probe substrates for UGT1A4, UGT1A6, and UGT1A9, respectively (Court, 2005). In this study, formation of trifluoperazine glucuronide, serotonin glucuronide, and mycophenolic acid phenolic glucuronide was used as an index reaction for UGT1A4, UGT1A6, and UGT1A9 enzymatic activities, respectively.
Materials and Methods
Chemicals and Reagents.
Trifluoperazine (TFP) (≥99%), serotonin (≥98%), potassium phosphate dibasic, Tris-HCl, UDP-glucuronic acid (UDPGA), β-glucuronidase, magnesium chloride, bovine serum albumin (BSA), alamethicin, niflumic acid, and epigallocatechin gallate (EGCG) (≥97%) were purchased from Sigma-Aldrich (St. Louis, MO). Acetonitrile, ethanol, methanol, acetone, hecogenin acetate (93%), and 1-naphthol (>99%) were purchased from Thermo Fisher Scientific (Waltham, MA). Serotonin-O-β-d-glucuronide was provided by RTI International (Research Triangle Park, NC) through the National Institute of Mental Health Chemical Synthesis Program. Mycophenolic acid (MPA) (98%), mycophenolic acid β-d-glucuronide (MPAG) (98%), and mycophenolic acid-d3-β-d-glucuronide (MPA-d3-G) (98%) were purchased from Toronto Research Chemicals (North York, ON, Canada). Herbal extracts (black cohosh, Cimicifuga racemosa; cranberry, Vaccinium marocarpon, echinacea, Echinacea purpurea; garlic, Allium sativum; ginkgo, Ginkgo biloba; ginseng, Panax ginseng; milk thistle, Silybum marianum; saw palmetto, Serenoa repens; and valerian, Valeriana officinalis) were generously provided by Finzelberg and Co. KG (Andernach, Germany) as dry powder. Table 1 summarizes the properties of the extracts screened. UltraPool human liver microsomes (HLM), which are microsomes pooled from 150 donors to minimize lot-to-lot variability, were purchased from BD Biosciences Discovery Labware (Bedford, MA).
List of herbal extracts screened for UGT1A4, UGT1A6, and UGT1A9 inhibition
Preparation of Herbal Working Solutions.
Herbal extracts were reconstituted with the solvents originally used for extraction and standardization by the vendor (Table 1). To remove any insoluble contents, the mixture was centrifuged at 20,000g for 5 min, and the liquid supernatant was removed. Working solutions were freshly prepared so that final herbal concentrations in screening incubations would represent the recommended daily intake of each extract in 53, 5.3, and 0.53 liters. These volumes roughly represent total body fluids, and two extremes of a range of concentrations that could appear in the small intestine, assuming 100% bioavailability as described previously by Hellum et al. (2007). For confirmation experiments, a range of concentrations around the rough IC50 of herbal extracts was used in incubations. The concentration of organic solvents in incubations was the same in all incubations including controls and was limited to 1%. For EGCG, working solutions were freshly prepared in 10% methanol and 1.5 mM ascorbic acid, which was added to ensure EGCG stability during the experiment (Lu et al., 2003). Ginseng and ginkgo were tested as both unhydrolyzed and acid-hydrolyzed extracts due to the high content of glycosides, which are extensively hydrolyzed in vivo by stomach acids and intestinal bacteria. Acid-hydrolyzed ginseng extract was prepared by dissolving 60 mg of the powder extract in 1 ml of 60% ethanol-40% 0.5 M HCl (Sloley et al., 2006). After 90 min at 37°C, the extract was neutralized with 0.1 M KOH and was serially diluted to prepare working solutions containing 10% ethanol. Acid-hydrolyzed ginkgo extract was prepared by dissolving 30 mg of the powder extract in 1 ml of 60% acetone-40% 5 M HCl. The acid-treated extract was heated at 90°C for 1 h and neutralized with 2 M KOH. Working solutions were prepared so that their concentrations were 10-fold higher than the final concentrations in incubations.
Incubations of Herbal Extracts with TFP.
TFP was used as a probe substrate for UGT1A4 in HLM. Incubations with TFP were performed as described previously by Uchaipichat et al. (2006). In brief, the incubation mixture (final volume, 250 μl) consisted of TFP, 5 mM MgCl2, 50 mM Tris-HCl buffer (pH 7.4), 0.1 mg/ml microsomal proteins, and alamethicin (100 μg/mg protein). The concentration of TFP in incubations was 60 μM, which corresponds to the Km in HLM (Uchaipichat et al., 2006). The mixture was preincubated on ice for 15 min. The reaction was started by adding UDPGA (final concentration, 5 mM). After the mixture was incubated for 20 min at 37°C, the reaction was stopped by adding 250 μl (4% acetic acid-96% methanol), vortex-mixing, and placing tubes on ice. Tubes were centrifuged for 10 min at 20,000g, and the supernatant was transferred to HPLC tubes. Screening experiments were performed by adding herbal extracts at three different concentrations to the incubation mixture. Incubations with and without hecogenin (50 μM) were performed to serve as positive and negative controls, respectively.
Chromatographic Analysis of TFPG.
HPLC analysis was performed with a Shimadzu LC-10AD VP pump (Shimadzu Scientific Instruments, Columbia, MD) connected to a Waters 717 autosampler and Waters 2475 florescence detector (Waters, Milford, MA). Fifty microliters of the incubation supernatant was injected on a reverse-phase Luna Phenyl-Hexyl column (2 × 100 mm, 3 μm; Phenomenex, Torrance, CA). Isocratic chromatography was carried out at ambient temperature using a mobile phase consisting of 0.1% trifluoroacetic acid in acetonitrile-deionized water (30:70) at a flow rate of 0.2 ml/min. The total run time was 15 min. TFPG was detected at an excitation wavelength of 310 nm and emission wavelength of 475 nm (Rele et al., 2004).
The identity of the TFPG peak was verified through enzymatic hydrolysis using β-glucuronidase. TFP (60 μM) was incubated with HLM as described above for 1 h at 37°C. Then, 25 μl of 100 mM potassium phosphate buffer (pH 4.0) and 2500 units of β-glucuronidase were added. Tubes were incubated for 16 h at 37°C. The reaction was stopped by adding 10 μl of 70% HClO4, vortex-mixing, and centrifugation at 20,000g for 10 min. The supernatant was transferred to HPLC tubes for injection. Control incubations were performed in the same way but did not contain β-glucuronidase enzyme. The TFPG peak was detected in the control incubation but not in the hydrolyzed incubation.
Incubations of Serotonin with Herbal Extracts.
To investigate the effect of herbal products on UGT1A6 activity, incubations of herbal extracts with HLM were performed using serotonin as a probe substrate as described by Krishnaswamy et al. (2003) with modifications. In brief, the incubation mixture (final volume, 100 μl) consisted of serotonin at a concentration at approximately the Km value in HLM (8 mM), 5 mM MgCl2, 50 mM Tris-HCl buffer (pH 7.4), 0.5 mg/ml microsomal proteins, and alamethicin (100 μg/mg protein). The mixture was preincubated on ice for 15 min. The reaction was started by adding UDPGA (final concentration, 5 mM). After the mixture was incubated for 60 min at 37°C, the reaction was stopped by adding 10 μl of 24% perchloric acid-acetonitrile (1:1, v/v), vortex-mixing, and placing tubes on ice. Tubes were centrifuged for 10 min at 20,000g, and the supernatant was transferred to HPLC tubes. 1-Naphthol (50 μM) was used as a positive control inhibitor in the screening assays (Fujiwara et al., 2008).
Chromatographic Analysis of Serotonin Glucuronide.
Isocratic chromatography was carried out at ambient temperature on a reverse-phase C18 Symmetry column (3.9 × 150 mm, 5 μm; Waters). The mobile phase consisted of 5% acetonitrile-95% 2 mM ammonium acetate (pH 2.7). Isocratic elution at a flow rate of 1.0 ml/min was used. The total run time was 10 min, and the injection volume was 30 μl. The HPLC system consisted of a Shimadzu LC-10AD VP pump connected to a Waters 717 autosampler and Waters 2475 florescence detector. Serotonin glucuronide was detected at an excitation wavelength of 225 nm and emission wavelength of 330 nm. To confirm the identity of the serotonin glucuronide peak, the retention time was compared with the serotonin glucuronide standard. In addition, the serotonin glucuronide peak was collected from the HPLC eluate and analyzed using MS/MS. The isolated fraction showed abundant ions with m/z 353, which matches the m/z of serotonin glucuronide ions in the positive mode. Upon fragmentation of the parent ion, a product ion with m/z 177 was produced, which matches the expected breakdown of the conjugate into glucuronic acid and free serotonin.
Incubations of MPA with Herbal Extracts.
Incubations with MPA were performed as described previously with modifications (Mohamed et al., 2008). In brief, the incubation mixture (100 μl) contained HLM (protein concentration, 0.16 mg/ml), alamethicin (100 μg/mg protein), MgCl2 (5 mM), 2% BSA, and 100 mM phosphate buffer, pH 7.4. MPA was used at a concentration equivalent to the Km value in HLM (240 μM). Microsomes were preincubated on ice with alamethicin for 15 min. The reaction was started by adding UDPGA (1 mM) and placing incubation tubes in a water bath at 37°C for 30 min. The reaction was stopped by adding 300 μl of ice-cold acetonitrile and 20 μl of internal standard (20 μg/ml MPA-d3-G). Tubes were vortex-mixed for 2 min and centrifuged for 10 min at 20,000g. The supernatant was diluted 12-fold with purified water, and 5 μl was injected into the HPLC system. Incubations of MPA with niflumic acid (70 μM) were used as positive controls (Vietri et al., 2000).
MPAG Liquid Chromatography-Tandem Mass Spectrometry Assay.
MPAG was determined by liquid chromatography-tandem mass spectrometry on a Thermo Finnigan Surveyor series HPLC system connected to a TSQ Quantum triple quadrupole mass spectrometer (Thermo Fisher Scientific) using electrospray ionization, as described previously (Mohamed et al., 2008). In brief, 5 μl of each sample was injected on a reverse-phase Synergi Fusion-RP18 column (100 × 2 mm, 4 μm; Phenomenex). The mobile phase consisted of (A) 1 mM acetic acid in deionized water and (B) 1 mM acetic acid in acetonitrile. Gradient elution at a flow rate of 0.22 ml/min was used with the following steps: at the start of the run, 30% B for 1 min, then increased to 90% B in 0.75 min, and held at 90% B between 1.75 and 3.1 min, and from 3.6 to 6.5 min, the column was reequilibrated at 30% B. Analysis was performed in the single reaction monitoring, negative ion mode using the mass transitions of m/z 495 → 319 and m/z 498 → 322 for MPAG and MPA-d3-G, respectively.
Data Analysis.
The remaining enzyme activity was calculated from the peak area of the glucuronide metabolites formed in herbal extract incubations expressed as a percentage of control. Remaining enzyme activity and herbal extract concentration data were fitted to eq. 1 using Prism 5.02 (GraphPad Software Inc., San Diego, CA) to estimate IC50 values.
Volume per dose index (VDI) was calculated using eq. 1 and was used as a measure of the potential of IC50 concentrations to be reached in vivo as described by Strandell et al. (2004). The VDI is defined as the volume in which one dose should be dissolved to obtain the corresponding IC50 concentration:
 where RDI is recommended daily intake.
where RDI is recommended daily intake.
Results
A total of 35 herb-UGT enzyme pairs were evaluated, each at three different concentrations. Results from the screening experiments are summarized in Table 2. Rough IC50 and VDI values were estimated on the basis of the remaining enzyme activity data at the three concentrations of each herbal extract. VDI was used to select the herb-UGT interactions to investigate further. A VDI cutoff value was considered to be 5 liters for UGT1A4 interactions and 2 liters for UGT1A6 and UGT1A9 interactions. This value was based on an expression study that showed that UGT1A6 and UGT1A9 are expressed in the intestine and the liver, whereas UGT1A4 is mainly expressed in the liver (Ohno and Nakajin, 2009). Herbal extracts that showed inhibition of a UGT enzyme with VDI values exceeding the specific cutoff value in the screening experiments were studied further in confirmatory assays to estimate accurate IC50 and VDI values. For all reported IC50 values, goodness of fit (r2) of the nonlinear regression curve was greater than 0.9.
Effect of commonly used herbal extracts on UGT1A4, UGT1A6, and UGT1A9 activity
Each herbal extract was coincubated at three concentrations with TFP (for UGT1A4), serotonin (for UGT1A6), and MPA (for UGT1A9) and HLM. Formation of TFPG, serotonin glucuronide, and MPAG were used as index reactions for activity of UGT1A4, UGT1A6, and UGT1A9 enzyme activities, respectively. Formation of glucuronides was compared in incubations with herbal extract with negative control incubations. Data represent best-fit IC50 values ± S.E. The goodness-of-fit r2 value was >0.9 for all reported IC50 values. VDI was calculated by dividing the daily intake of each herb by the rough IC50 value. RDI values were determined on the basis of the PDR for Herbal Medicines (Gruenwald et al., 2004) and commercially available products.
Effect of Herbal Extracts on TFPG Formation.
The effect of 10 herbal extracts on UGT1A4 activity was achieved through incubations of pooled HLM with TFP and monitoring formation of TFPG as an index of UGT1A4 activity. For milk thistle and acid-hydrolyzed ginkgo extracts, evaluation of their effects on UGT1A4 activity was not possible because of interference of the herbal extracts with TFPG fluorescence. All the tested extracts inhibited TFPG formation with different potencies (Fig. 1). Herbal extracts showing rough IC50 values less than 100 μg/ml were (mean ± S.E.) EGCG (34.4 ± 4.1 μg/ml), black cohosh (69.7 ± 4.8 μg/ml), and saw palmetto (70.6 ± 9.3 μg/ml) (Table 2). Only EGCG inhibited UGT1A4 with a VDI value exceeding 5 liters. This finding was confirmed by incubating TFP with increasing concentrations of EGCG. Best-fit IC50 was 33.8 ± 3.1 μg/ml and VDI value was 7.4 liters on the basis of the daily dose of 250 mg (Table 3; Fig. 2).

Effect of commonly used herbal extracts on UGT1A4, UGT1A6, and UGT1A9 enzyme activities. HLM were coincubated with herbal extracts and TFP for UGT1A4 activity (A), serotonin for UGT1A6 activity (B), and mycophenolic acid for UGT1A9 activity (C). Three concentrations were tested for each herbal extract, which represent extract daily intake in 53 liters (dotted bars), 5.3 liters (checkered bars), and 0.53 liters (striped bars). Formation of TFPG, serotonin glucuronide, and MPAG was detected in the respective herbal incubations. Percentage of activity was calculated as the percentage of the glucuronide peak area in herbal incubations compared with that in negative controls. Each value represents mean of duplicate incubations. Error bars represent positive S.E.
Determination of inhibitory potency of selected UGT1A4, UGT1A6, and UGT1A9 herbal inhibitors
Extracts were selected if their VDI on the basis of rough IC50 values exceeded 4 liters for UGT1A4 or 2 liters for UGT1A6 and UGT1A9. Several concentrations of each extract were coincubated with alamethicin-activated HLM and TFP (for UGT1A4), serotonin (for UGT1A6), or MPA (for UGT1A9). The percentage of remaining activity was measured as the formation of each glucuronide in herbal incubation as a percentage of the negative control. IC50 values were calculated by fitting data points to the Hill IC50 equation as described under Materials and Methods. Values reported are best-fit IC50 values ± S.E. The goodness-of-fit r2 value was ≥0.95 for all reported IC50 values. VDI was calculated by dividing the daily intake of each herb by the IC50 value. RDI values were determined on the basis of the PDR for Herbal Medicines (Gruenwald et al., 2004) and commercially available products.

Inhibitory effect of green tea catechin EGCG on TFPG formation in HLM. Increasing concentrations of EGCG were incubated with 60 μM TFP, 0.1 mg/ml alamethicin-activated HLM, 5 mM UDPGA, and 5 mM MgCl2 for 20 min at 37°C. Formation of TFPG was used as an index for UGT1A4 activity in HLM incubations. Each data point represents the mean of duplicate incubations. Error bars represent two-sided S.E.M. Data points were fitted to the IC50 equation as described under Materials and Methods. The goodness-of-fit r2 value was 0.98.
Effect of Herbal Extracts on Serotonin Glucuronide Formation.
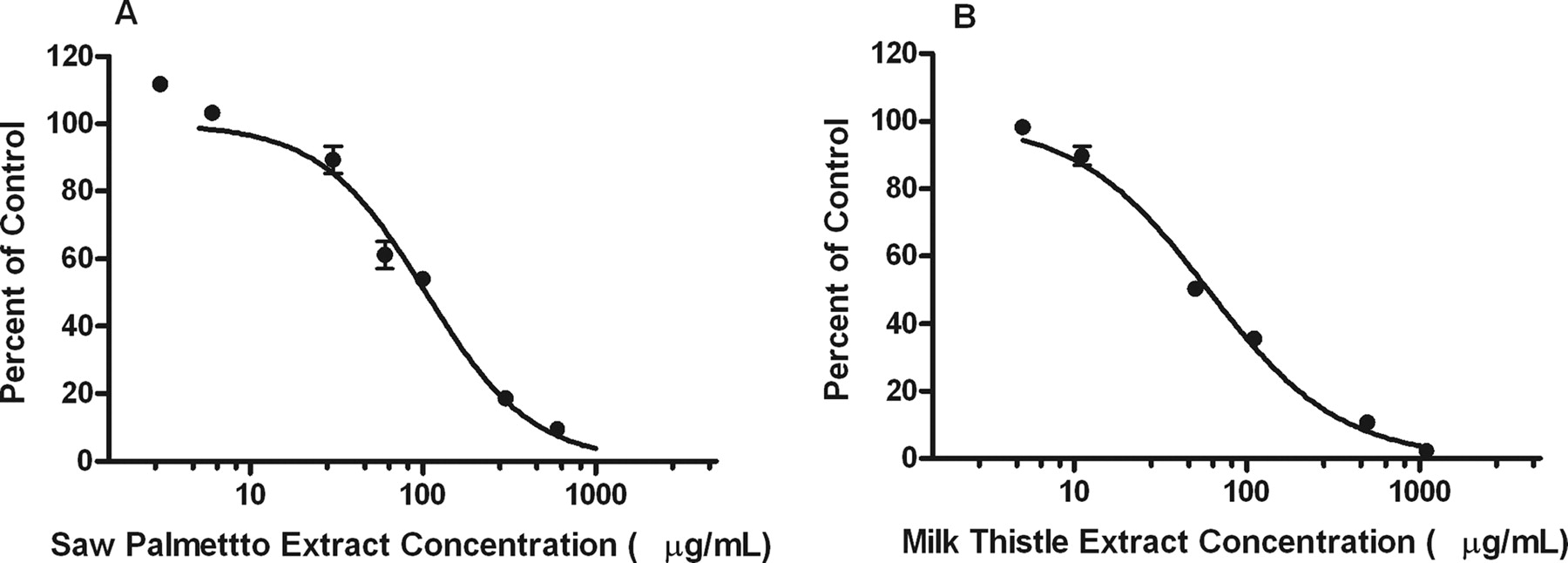
Milk thistle, saw palmetto, EGCG, and echinacea inhibited serotonin glucuronide formation with IC50 values of 66.9 ± 3.5, 131.8 ± 21.5, and 183.6 ± 29.8 μg/ml, respectively (Table 2; Fig. 1). A VDI cutoff value of 2 liters was applied to select which extracts to study further. Only saw palmetto and milk thistle exceeded the VDI cutoff with values of 2.4 liters and 9.0, respectively (Table 2).
Precise IC50 and VDI values were determined for inhibition of serotonin glucuronide formation by milk thistle and saw palmetto (Fig. 3). Best-fit IC50 values were 59.5 ± 3.6 and 103.5 ± 10.7 for milk thistle and saw palmetto, respectively. VDI values were 6.3 and 3.1 liters for milk thistle and saw palmetto, respectively (Table 3).

Inhibition of serotonin glucuronide formation by saw palmetto and milk thistle extracts. Increasing concentrations of saw palmetto (A) and milk thistle (B) extracts were incubated with 8 mM serotonin, 5 mM MgCl2, 0.5 mg/ml alamethicin-activated HLM, and 5 mM UDPGA for 60 min at 37°C. Serotonin glucuronide formation was used as an index of UGT1A6 enzyme activity in HLM incubations. Each data point represents the mean of duplicate incubations. Error bars represent two-sided S.E.M. Data points were fitted to IC50 equation as described under Materials and Methods. The goodness-of-fit r2 value was 0.96 and 0.99 for saw palmetto and milk thistle, respectively.
Effect of Herbal Extracts on MPAG Formation.
Black cohosh, cranberry, echinacea, ginseng, acid-hydrolyzed ginseng, and milk thistle inhibited MPAG formation (Fig. 1). However, only milk thistle (rough IC50 = 35.9 ± 4.3 μg/ml; VDI = 16.7 liters) and cranberry (rough IC50 = 260.5 ± 33.0 μg/ml; VDI = 3.8 liters) exceeded the VDI cutoff of 2 liters and were selected for further study (Table 2). Precise best-fit IC50 and VDI values for milk thistle and cranberry were 33.6 ± 3.1 μg/ml and 17.9 liters and 230.4 ± 32.9 μg/ml and 3.1 liters, respectively) (Table 3; Fig. 4).
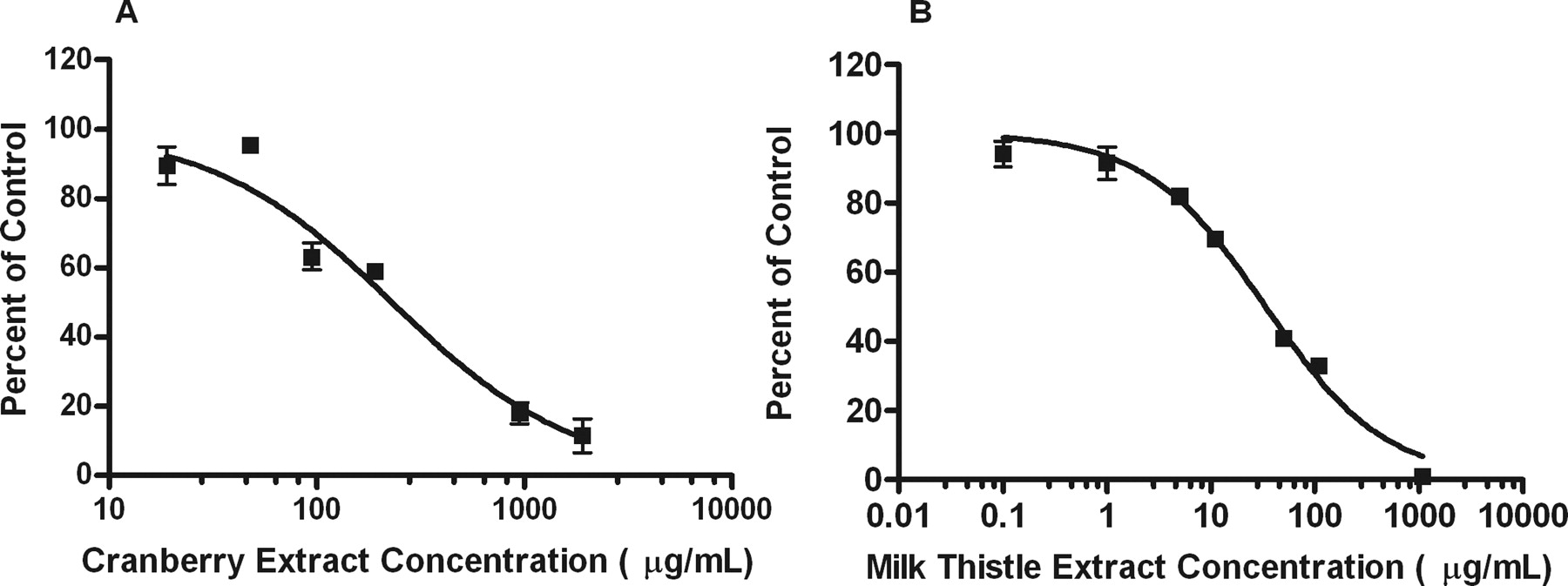
Inhibition of MPAG formation by cranberry and milk thistle extracts. Increasing concentrations of cranberry (A) and milk thistle (B) extracts were incubated with 240 μM mycophenolic acid, 5 mM MgCl2, 2% BSA, 0.16 mg/ml alamethicin-activated HLM, and 1 mM UDPGA. Formation of MPAG was used as an index for UGT1A9 activity in HLM incubations. Each data point represents the mean of duplicate incubations. Error bars represent two-sided S.E.M. Data points were fitted to the IC50 equation as described under Materials and Methods. The goodness-of-fit r2 value was 0.95 and 0.99 for cranberry and milk thistle, respectively.
Discussion
In this study, 10 commonly used herbal extracts were screened for their effects on the glucuronidation activity of UGT1A4, UGT1A6, and UGT1A9 in pooled HLM. UGT enzyme activities were measured in vitro using selective substrates: TFP for UGT1A4, serotonin for UGT1A6, and MPA for UGT1A9 (Court, 2005). On the basis of VDI values, the most potent inhibitors were EGCG for UGT1A4, milk thistle for both UGT1A6 and UGT1A9, saw palmetto for UGT1A6, and cranberry for UGT1A9. These findings highlight the possibility of herb-drug interactions through modulation of UGT enzyme activity. The likelihood of the observed in vitro interactions to occur in vivo depends on characteristics of the herb, the drug substrate, the specific enzyme, and the potency of the inhibition.
UGT1A4 is known to be the primary enzyme that catalyzes N-glucuronidation of primary, secondary, and aromatic amines, which include TFP, lamotrigine, tamoxifen, tacrolimus, and azole antifungals (Kiang et al., 2005; Rowland et al., 2006; Bourcier et al., 2010; Zhou et al., 2010; Laverdière et al., 2011). In addition, UGT1A4 shows O-glucuronidation activity toward steroidal compounds (Green and Tephly, 1996). Hecogenin is a known inhibitor of UGT1A4-mediated TFP glucuronidation with IC50 values of 1.5 μM (Uchaipichat et al., 2006). Compared with hecogenin, EGCG is a nonselective UGT1A4 inhibitor with moderate potency. EGCG has previously been shown to inhibit estradiol-3-O-glucuronidation, an index for UGT1A1 activity, with a lower IC50 value (7.8 μg/ml) (Mohamed et al., 2010). In addition, in this study EGCG showed some weak inhibitory activities toward UGT1A6 and UGT1A9 (Fig. 1). Pharmacokinetic studies show that maximum plasma concentrations of EGCG are more than 10-fold less than the observed IC50 values after consumption of high-dose (800 mg) EGCG (Foster et al., 2007). This finding suggests that inhibition of UGT1A4-mediated systemic glucuronidation by EGCG is unlikely. However, on the basis of VDI of the inhibition of 7.4 liters for a 250-mg dose, an effect of EGCG on hepatic first-pass metabolism of UGT1A4 substrates is possible and will be augmented with higher EGCG doses. EGCG has been studied at doses that reach 800 mg daily for its antioxidant and anticancer effects (Chow et al., 2005). With higher doses of EGCG (800 mg), the VDI will be 23.6 liter, indicating that the 800-mg dose can be diluted in up to 23.6 liters and still inhibit UGT1A4 activity by up to 50%. The effect of EGCG on glucuronidation of the UGT1A4 substrates TFP, lamotrigine, tamoxifen, and imipramine warrants further investigation.
UGT1A6 is typically a low-affinity enzyme that catalyzes glucuronidation of drug substrates including acetaminophen, naproxen, and deferiprone (Bowalgaha et al., 2005; Kiang et al., 2005; Limenta et al., 2008). Milk thistle and saw palmetto inhibited serotonin glucuronidation with IC50 concentrations attainable if the daily doses of milk thistle (600 mg) or saw palmetto (320 mg) are diluted with 6.3 and 3.1 liters, respectively. The observed milk thistle IC50 for UGT1A6 is equivalent to a total flavonolignan concentration of 22.6 μg/ml; this is approximately 1000-fold higher than the observed plasma concentration after intake of 600 mg of milk thistle extract (Schrieber et al., 2008). Taken together, these results indicate that milk thistle extract is more likely to inhibit UGT1A6-mediated first-pass rather than systemic metabolism. On the other hand, no pharmacokinetic data on saw palmetto are available. On the basis of an IC50 value exceeding 100 μg/ml and VDI of 3.1 liters, saw palmetto will be expected to have mild, if any, inhibition of UGT1A6-mediated metabolism in vivo (Table 2).
UGT1A9 catalyzes glucuronidation of a wide range of substrates including MPA, propofol, raloxifene, and flavopiridol (Kiang et al., 2005). In the current study, milk thistle and cranberry inhibited MPAG formation, which was used as an index reaction for UGT1A9 activity in HLM (Court, 2005). For milk thistle extract, the IC50 value was 33.6 μg/ml, which is equivalent to 12.7 μg/ml flavonolignans. Again, this concentration is much higher than the expected plasma concentration of flavonolignans after milk thistle intake (Schrieber et al., 2008). Therefore, inhibition of systemic metabolism of UGT1A9 substrates by milk thistle extract is not likely. Conversely, on the basis of the range of intestinal fluid volume of 0.5 to 5 liters, a single 600-mg dose of milk thistle may result in putative concentrations of 120 to 1200 μg/ml. Thus, inhibition of first-pass metabolism of UGT1A9 substrates by milk thistle extract is possible.
In this study, we screened specific UGT enzyme activities using HLM rather than human intestine microsomes (HIM) or expressed enzymes. The difference between expressed enzymes and HLM is that the first contain single UGT enzymes, whereas the latter contain all the hepatic isoforms. Therefore, HLM are closer to the in vivo environment because of the availability of other UGT enzymes that may form heterodimers, which has been reported for some UGT enzymes and may affect enzyme activity (Ouzzine et al., 2003). Because our goal was to screen for interactions that may have clinical significance, the use of HLM was more appropriate. This was made feasible by the availability of selective substrates for different UGT enzymes in HLM (Court, 2005). Likewise, HIM contain all the intestinal UGT enzymes. However, no selective substrates for individual UGT enzymes have been described in HIM.
Calculation of VDI provides a helpful tool to predict the likelihood of achieving IC50-equivalent concentrations in the intestine or plasma in the absence of clinical data (Strandell et al., 2004). Although this approach is sufficient for the purpose of screening and hypothesis generation, it is limited by not considering the extent of absorption of phytochemicals through tissue and cellular barriers. Use of VDI assumes that the concentration in the gastrointestinal lumen is equivalent to that in the endoplasmic reticulum of intestinal epithelial cells where UGT enzymes are located. This assumption may lead to overestimation of the extent of the inhibition, because many phytochemicals are poorly absorbed through the intestinal wall. Therefore, the results need to be confirmed in clinical studies and, where available, IC50 values need to be compared with unbound in vivo concentrations. It is worth noting that using VDI to describe inhibition potency changes the order of significance of inhibitors compared with IC50 values. For example, based on rough IC50 values, black cohosh and saw palmetto are equipotent inhibitors of UGT1A4 activity (rough IC50 = 69.7 and 70.6 μg/ml) (Table 2). However, the daily dose of saw palmetto is 8-fold higher than that of black cohosh (320 versus 40 mg). Thus, ingestion of 320 mg of saw palmetto is expected to result in a higher extent of UGT1A4 inhibition than ingestion of 60 mg of black cohosh.
In summary, in this study, 10 herbal extracts were screened for inhibition of three UGT1A enzymes: UGT1A4, UGT1A6, and UGT1A9. We report inhibition of UGT1A4 by EGCG, UGT1A6 by milk thistle and saw palmetto, and UGT1A9 by cranberry and milk thistle extracts. Among these, EGCG inhibition of UGT1A4 and milk thistle inhibition of UGT1A6 and UGT1A9 are likely to affect first-pass glucuronidation of substrates, which will be important for drugs with high first-pass extraction ratios. The in vivo effects of these interactions on the pharmacokinetics of UGT1A4, UGT1A6, and UGT1A9 substrates remain to be determined in clinical studies.
Authorship Contributions
Participated in research design: Mohamed and Frye.
Conducted experiments: Mohamed.
Performed data analysis: Mohamed.
Wrote or contributed to the writing of the manuscript: Mohamed and Frye.
Acknowledgments
We thank Dr. Veronika Butterweck and Finzelberg and Co. KG (Andernach, Germany) for providing the herbal extracts.
Footnotes
This work was supported in part by the National Institutes of Health National Center for Complementary and Alternative Medicine [Grant R21-AT005083].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.111.039602.
-
ABBREVIATIONS:
- UGT
- UDP-glucuronosyltransferase
- TFP
- trifluoperazine
- UDPGA
- UDP-glucuronic acid
- BSA
- bovine serum albumin
- EGCG
- epigallocatechin gallate
- MPA
- mycophenolic acid
- MPAG
- mycophenolic acid-7-O-glucuronide
- MPA-d3-G
- mycophenolic acid-d3-β-d-glucuronide
- HLM
- human liver microsome(s)
- HPLC
- high-performance liquid chromatography
- TFPG
- TFP glucuronide
- VDI
- volume per dose index
- HIM
- human intestinal microsomes.
- Received March 16, 2011.
- Accepted June 1, 2011.
- Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}