


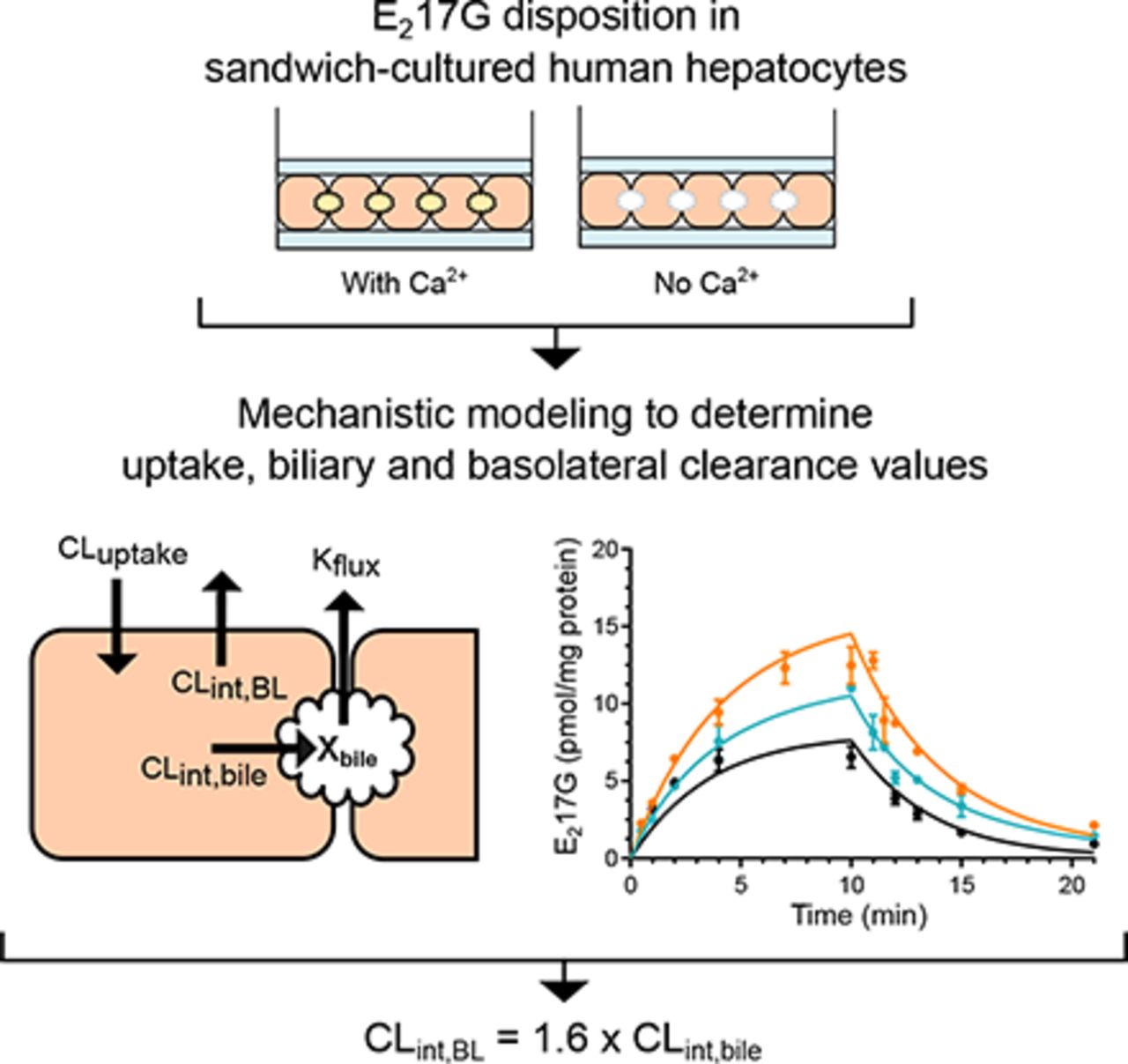
Visual Overview

Abstract
Estradiol-17β-glucuronide (E217G) is an estrogen metabolite that has cholestatic properties. In humans, circulating E217G is transported into hepatocytes by organic anion transporting polypeptides (OATPs) and is excreted into bile by multidrug-resistance associated protein 2 (MRP2). E217G is also a substrate of the basolateral efflux transporters MRP3 and MRP4, which translocate E217G from hepatocytes to blood. However, the contribution of basolateral efflux to hepatocyte disposition of E217G has not been evaluated previously. To address this question, E217G disposition was studied in sandwich-cultured human hepatocytes and mechanistic modeling was applied to calculate clearance values (mean ± S.D.) for uptake, intrinsic biliary excretion (CLint,bile) and intrinsic basolateral efflux (CLint,BL). The biliary excretion index of E217G was 45% ± 6%. The CLint,BL of E217G [0.18 ± 0.03 (ml/min)/g liver) was 1.6-fold higher than CLint,bile [0.11 ± 0.06 (ml/min)/g liver]. Simulations were performed to study the effects of increased CLint,BL and a concomitant decrease in CLint,bile on hepatic E217G exposure. Results demonstrated that increased CLint,BL can effectively reduce hepatocellular and biliary exposure to this potent cholestatic agent. Simulations also revealed that basolateral efflux can compensate for impaired biliary excretion and, vice versa, to avoid accumulation of E217G in hepatocytes. However, when both clearance processes are impaired by 90%, hepatocyte E217G exposure increases up to 10-fold. These data highlight the contribution of basolateral efflux transport, in addition to MRP2-mediated biliary excretion, to E217G disposition in human hepatocytes. This elimination route could be important, especially in cases where basolateral efflux is induced, such as cholestasis.
SIGNIFICANCE STATEMENT The disposition of the cholestatic estrogen metabolite estradiol-17β-glucuronide (E217G) was characterized in sandwich-cultured human hepatocytes. The intrinsic basolateral efflux clearance was estimated to be 1.6-fold higher than the intrinsic biliary excretion clearance, emphasizing the contribution of basolateral elimination in addition to biliary excretion. Simulations highlight how hepatocytes can effectively cope with increased E217G through the regulation of both basolateral and biliary transporters.
Introduction
Estradiol-17β-glucuronide (E217G) is a metabolite of estradiol, an estrogen steroid hormone. E217G is formed by uridine 5′-diphospho-glucuronosyltransferase-mediated glucuronidation of estradiol, primarily in the liver (Musey et al., 1997; Itäaho et al., 2008). Hepatic concentrations of E217G are regulated by multiple transport proteins expressed in the liver. E217G is a well-known substrate of multidrug-resistance associated protein 2 (MRP2, ABCC2), which mediates E217G excretion into bile (Morikawa et al., 2000). On the basolateral (sinusoidal) membrane, organic anion transporting polypeptides 1B1 and 1B3 (OATP1B1, SLCO1B1 and OATP1B3, SLCO1B1) mediate the uptake of E217G into hepatocytes from the blood (Karlgren et al., 2012). In fact, E217G is commonly used as a probe substrate in OATP1B1/1B3 and MRP2 inhibition studies in vitro (Pedersen et al., 2008; Karlgren et al., 2012; Brouwer et al., 2013; Morgan et al., 2013). In addition, E217G is transported by MRP3 (ABCC3) and MRP4 (ABCC4), which pump substrates from hepatocytes into sinusoidal blood on the basolateral membrane (Zeng et al., 2000; Chen et al., 2001). However, the contribution of basolateral efflux to the hepatic disposition of E217G has not been elucidated.
E217G transport is of interest because it is a potent cholestatic agent and it is postulated to be involved in the pathogenesis of intrahepatic cholestasis of pregnancy (ICP). ICP is a condition that is characterized by fairly harmless pruritus and elevated serum bile acid concentrations in the mother, but can increase the risk for preterm labor or even intrauterine death (Geenes and Williamson, 2009). Maternal symptoms typically present in the third trimester of pregnancy, coinciding with the highest estrogen levels during pregnancy. Therefore, it is thought that elevated hepatic E217G concentrations may play a role in triggering ICP. Understanding the mechanisms underlying the hepatic disposition of E217G is important from the viewpoint of preventing and managing ICP.
Most of the studies regarding the cholestatic effects of E217G have been performed in rats. E217G administration (2 µmol infused over 18 minutes) decreased bile flow by 51% in perfused rat livers (Huang et al., 2000). Additionally, E217G decreased the elimination of the hepatobiliary imaging agent indocyanine green in female rhesus monkeys (Slikker et al., 1983). MRP2 seems to play a key role in E217G-induced cholestasis, because even doses 12-fold higher than the dose required for cholestatic effects in wild-type rats did not decrease bile flow in Mrp2-deficient TR− rats (Huang et al., 2000). Studies in Mrp2-deficient rats have shown that Mrp2 mediates the secretion of glutathione into bile and that this is important for bile acid-independent bile flow (Takikawa et al., 1991; Elferink and Groen, 2002). Overall, E217G appears to exert its cholestatic effects through several mechanisms. Administration of E217G to rats increased the internalization of Mrp2 and the bile salt export pump (Bsep) from the cell membrane by clathrin-mediated endocytosis (Mottino et al., 2002; Miszczuk et al., 2018). In addition, E217G increased the accumulation of bile acids through trans-inhibition of BSEP (Stieger et al., 2000; Vallejo et al., 2006). An increase in intrahepatic bile acid concentrations could induce organic solute transporter α/β expression (Boyer et al., 2006) and enhance bile acid efflux into the blood, which might explain the observed increase in serum bile acids in ICP (Walker et al., 2002).
Although the current literature suggests a possible role for MRP2 in E217G-induced ICP, as discussed above, several mechanisms appear to be involved in the development of ICP, and MRP2 is not the only process regulating hepatic E217G exposure. The exposure also is affected by hepatic basolateral uptake and efflux transporters. Therefore, in this study, the contribution of basolateral efflux in the hepatic disposition of E217G was quantitatively evaluated by uptake and efflux studies using sandwich-cultured human hepatocytes (SCHH) and mechanistic modeling. The clearance estimates derived from the model were used to simulate conditions resulting in inhibition or induction of the different pathways affected by cholestasis or drug-drug interactions based on previously published literature. The results of this study reinforce the notion that basolateral efflux can act as an important excretion route for E217G, especially in cholestatic conditions where basolateral transporters (e.g., MRP3, MRP4) are upregulated.
Materials and Methods
Materials.
Transporter certified cryopreserved human hepatocytes (three female donors, Table 1) were obtained from ThermoFisher Scientific (Waltham, MA) and In Vitro ADMET Laboratories (Columbia, MD). QualGro™ seeding medium and QualGro™ hepatocyte culture medium were obtained from BioIVT (Durham, NC). Tritium-labeled estradiol-17β-glucuronide ([3H]-E217G) (52.9 Ci/mmol, radiochemical purity >97%) and taurocholate ([3H]-TCA) (9.7 Ci/mmol, radiochemical purity >97%) were purchased from PerkinElmer (Boston, MA). Nonlabeled E217G and TCA were purchased from Sigma-Aldrich (St. Louis, MO). Standard Hank’s balanced salt solution (HBSS) and HBSS without calcium and magnesium were purchased from ThermoFisher Scientific. Collagen (type I)-coated BioCoat™ 24-well cell culture plates and Matrigel® were obtained from BD Biosciences (San Jose, CA).
Donor information for the three hepatocyte lots used in the studies
All donors were women.
Sandwich-Cultured Human Hepatocytes.
On day 0, human hepatocytes from the three donors were seeded on BioCoat™ 24-well plates at a density of 0.45 × 106 hepatocytes per well using QualGro™ seeding medium and overlaid with Matrigel® diluted in QualGro™ hepatocyte culture medium (0.25 mg/ml) on day 1. SCHH were maintained in QualGro™ hepatocyte culture medium, which was changed daily until experiments were performed on day 5.
Disposition of E217G in SCHH.
The uptake and efflux studies were conducted by applying B-CLEAR© technology (BioIVT) according to the schemes shown in Fig. 1 (Liu et al., 1999). Briefly, SCHH were washed twice and preincubated for 10 minutes at 37°C with either standard HBSS (containing calcium and magnesium) or Ca2+-free HBSS (HBSS without calcium and magnesium, supplemented with 1 mM EGTA). After preincubation, the uptake phase was initiated by incubating SCHH with 0.3 μM [3H]-E217G (2 μCi/ml) in standard HBSS at 37°C. E217G accumulation in cells and biliary networks (cells + bile; measured in SCHH preincubated with standard HBSS) and E217G accumulation in cells (measured in SCHH preincubated with Ca2+-free HBSS) was determined during the uptake phase by terminal sampling at designated time points. After the 10-minute uptake phase, buffers containing E217G were removed from the wells and cells were washed three times with 37°C standard or Ca2+-free HBSS buffer followed by initiation of the efflux phase. During the efflux phase, E217G efflux into the buffer and the amount remaining in cells and biliary networks (standard HBSS preincubation) and cells (Ca2+-free HBSS preincubation) were determined by terminal sampling at designated time points. In addition, B-CLEAR© studies were performed as above with taurocholate (TCA) for comparison with the E217G results. In these studies, SCHH were incubated with 1 µM TCA (0.8 μCi/ml) for 10 minutes in standard HBSS after a 10-minute preincubation in standard HBSS or Ca2+-free HBSS. Accumulation of TCA in the cells and cells + bile was determined by terminal sampling.

Scheme depicting the uptake and efflux studies performed in SCHH with E217G. (A) Test compound (E217G) is taken up into the cells via uptake transporters (CLuptake) and excreted from the cell to the medium by transporters on the basolateral membrane (CLint,BL) or into the bile by canalicular transporters (CLint,bile). In the presence of Ca2+, E217G accumulates in the bile canaliculi but can flux into the medium (Kflux) with contractions of the canaliculi. When incubated in Ca2+-free buffer, the tight junctions of the bile canaliculi are disrupted and all excreted compound returns to the buffer. (B) Summary of the SCHH uptake and efflux study protocol performed in the presence and absence of Ca2+ to determine biliary excretion.
The radioactivity in samples from cells, cells + bile, and buffer was quantified by liquid scintillation counting using a Tri-Carb 3100TR counter (Perkin Elmer). Prior to analysis, cells were lysed with lysis solution containing 1× phosphate-buffered saline with 0.5% Triton X-100. Total protein concentration of the cell lysates was determined using the Pierce™ BCA Protein Assay Kit (ThermoFisher Scientific). The amount of E217G and TCA measured in the cell and the buffer samples was normalized to the protein amount in the wells. The biliary excretion index (BEI) was calculated from data at the end of the 10-minute uptake phase using eq. 1: (1)where Xcells+bile is the amount of test compound (E217G or TCA) accumulated in the cells and bile networks in standard HBSS, and Xcells is the amount of test compound accumulated in the cells that were preincubated in Ca2+-free HBSS.
(1)where Xcells+bile is the amount of test compound (E217G or TCA) accumulated in the cells and bile networks in standard HBSS, and Xcells is the amount of test compound accumulated in the cells that were preincubated in Ca2+-free HBSS.
Mechanistic Modeling of E217G in SCHH.
Mechanistic modeling was applied to the uptake and efflux data using Phoenix WinNonlin, v7.0 (Certara USA, Princeton, NJ) using a previously reported approach (Pfeifer et al., 2013). Briefly, the mechanistic model depicted in Fig. 1 was fit simultaneously to the E217G uptake and efflux in standard and Ca2+-free HBSS versus time data. The basolateral uptake clearance (CLuptake), intrinsic basolateral efflux clearance (CLint,BL), intrinsic biliary excretion clearance (CLint,bile), and the first-order rate constant for flux from the bile network into the buffer (Kflux) were determined. Assuming non-saturable clearances for all the pathways, the model equations are as follows:
Mass of E217G in standard HBSS (X+buffer):
 (2)
(2)Mass of E217G in Ca2+-free HBSS (X-buffer):
(3)Mass of E217G in cells (Xcells):
(4)Mass of E217G in bile (Xbile):
(5)Mass of E217G in cells + bile (Xcells+bile):
(6)





The first-order rate constant Kwash was fixed at 104 minute−1 between 10 and 11 minutes in the simulations. This parameter was included to remove all E217G from the buffer and mimic the wash step between the uptake and efflux phases (Pfeifer et al., 2013; Yang et al., 2015). The initial mass of E217G in both buffers was set equal to the dose, and the initial mass in the other compartments was set equal to zero. The in vitro observations used for data fitting were E217G Xcells+bile and X+buffer in standard HBSS, and Xcells and X−buffer in Ca2+-free conditions. An additive error model was used for the mass of E217G in standard HBSS and in cells incubated in Ca2+-free buffer, and a Poisson error model (a power model with a coefficient of 0.5) was used for the other observations. The initial estimate for CLuptake was calculated using the first three time points in the uptake phase as (dXcells+bile/dt)/Cbuffer. The total efflux clearance (CLint, BL + CLint, bile) was estimated from efflux phase data under Ca2+-free conditions, where (CLint, bile + CLint, BL) = X−Buffer,21 min/area under the curve (AUC)cells,11–21 min. Initial estimates were derived by dividing CLint, bile + CLint, BL by 2. The ordinary differential equations were solved using the matrix exponent ordinary differential equations solver in Phoenix WinNonlin. The goodness of fit was evaluated based on visual observation of predicted and observed E217G mass and the CV% of the derived parameter estimates. The model derived clearance values were scaled to the liver and expressed as milliliter per minute per gram liver, assuming a protein content of 90 mg protein/g liver (Sohlenius-Sternbeck, 2006). The fraction excreted by the two elimination pathways (febile and feefflux,BL) were calculated as the ratio of the specific clearance pathway divided by the sum of both clearance values (Zamek-Gliszczynski et al., 2009).
Effect of Transporter Induction and Inhibition.
The effect of transporter induction on E217G disposition was evaluated by simulating disposition in SCHH using the average clearance estimates derived from the mechanistic modeling. Simulations were performed using Stella version 9 (Isee Systems Inc., Lebanon, NH) and eqs. 2–6. CLint,bile was decreased and CLint,BL was increased by 2- and 4-fold in the simulations. These values were chosen to represent plausible changes in protein levels based on available literature (Thakkar et al., 2017). The BEI was calculated for each of the simulations using eq. 1 and simulated E217G amounts after 10-minute uptake. Simulations (up to 60 minutes) were performed to evaluate the changes in E217G in the cells and bile compartments at steady-state due to transporter induction. In addition, the effect of inhibition of CLint,bile and/or CLint,BL on E217G accumulation at steady state was calculated using eq. 7 and varying the clearance values: (7)where Kp is the hepatocyte to buffer concentration ratio. All data from the simulations and Kp calculations were normalized to control values (no inhibition or induction).
(7)where Kp is the hepatocyte to buffer concentration ratio. All data from the simulations and Kp calculations were normalized to control values (no inhibition or induction).
Results
All three hepatocyte lots showed time-dependent accumulation of E217G into cells during the uptake phase and excretion into the buffer during the efflux phase (Fig. 2). The biliary excretion index (BEI) of E217G after 10-minute uptake was 40%–52% in the three lots (Fig. 3). The functionality of the hepatocyte lots was verified with TCA; the BEI of TCA in the same hepatocyte lots was 41%–62%. All of the observed time course data for E217G uptake and efflux were described adequately by the model, and the CVs of the derived parameters were ≤68%. The largest variability in parameter estimates between lots was seen for CLint,bile (55% CV between lots). The average value of CLint,BL [0.18 ± 0.03 (ml/min)/g liver) was 1.6-fold higher than CLint,bile [0.11 ± 0.06 (ml/min)/g liver) (Table 2). The average fraction of E217G excreted via basolateral efflux (feefflux,BL) was 62%, and 38% was excreted via biliary excretion (febile).

E217G mass (pmol/mg protein) vs. time data in SCHH during the uptake and efflux phase. SCHH were incubated with 0.3 µM [3H]-E217G (2 µCi/ml) according to the scheme presented in Fig. 1B. The simulated mass-time profiles were generated using the relevant equations based on the model scheme depicted in Fig. 1A; the final parameter estimates are reported in Table 2. Data are presented as the mean ± S.D. (n = 3 wells, except n = 2 for cell lysate sample in Lot 1 at 0.5 minute with Ca2+-free HBSS).

Biliary excretion of E217G and TCA in SCHH. Accumulation (mean ± S.D.; n = 3) of substrate in SCHH after incubation with 0.3 µM [3H]-E217G (0.7 µCi/ml) or 1 µM [3H]-TCA (0.8 µCi/ml) for 10 minutes in standard HBSS after a 10-minute preincubation in standard HBSS (black bars; cells + bile) or Ca2+-free HBSS (white bars; cells). The BEI was calculated according to eq. 1.
Final parameter estimates from the model fitting
Sensitivity analysis revealed that hepatic E217G exposure is sensitive to CLint,BL; a 64% decrease in hepatic E217G exposure was observed when basolateral efflux was increased by fourfold (Fig. 4A). A decrease in CLint, bile caused an increase in hepatic E217G exposure, especially when CLint,BL was unchanged. However, when CLint,BL was simultaneously increased fourfold, even a fourfold reduction in CLint,bile had minimal effects on E217G in hepatocytes. E217G in bile was decreased to a similar extent by a twofold increase in CLint,BL and a twofold decrease in CLint, bile (Fig. 4B). The same was true for a fourfold increase in CLint,BL and a fourfold decrease in CLint,bile (Fig. 4B).

Effects of alterations in CLint,BL and CLint,bile on E217G disposition in SCHH. The impact of designated fold changes in clearance estimates on mean steady-state levels of E217G in cells (A) and bile (B), and on the BEI (C) at 10 minutes are plotted. All values were normalized to control simulations run using the average clearance estimates of E217G from the mechanistic modeling.
The maximal simulated decrease in biliary E217G was 90%, which was achieved with a fourfold increase in CLint,BL and a fourfold decrease in CLint,bile (Fig 4B). The BEI was only marginally affected by changes in CLint,BL; the BEI of E217G was decreased more than 30% when CLint,bile was reduced twofold and more than 60% when CLint,bile was reduced fourfold (Fig. 4C).
The results of simulations to evaluate the impact of inhibition of biliary excretion and/or basolateral efflux of E217G on hepatic exposure are plotted in Fig. 5. As shown, only modest changes (under twofold) in E217G hepatic exposure occurred when CLint,bile was inhibited by 90% because basolateral efflux compensated for impaired biliary excretion. Likewise, E217G hepatic exposure increased no more than 2.3-fold when CLint,BL was inhibited by 90% due to compensation by biliary excretion. However, up to a 10-fold increase in E217G hepatic exposure was observed when both CLint,BL and CLint,bile were inhibited simultaneously up to 90% of control values.

Effects of 0%–90% efflux inhibition on hepatic E217G exposure. The hepatocyte to buffer concentration ratio (Kp, eq. 7) was calculated using average clearance values (control) and decreasing either CLint,BL and/or CLint,bile. Exposure is shown as fold change from control (no inhibition). When CLint,bile is inhibited by 90%, hepatic E217G exposure is 1.5-fold (point A). When CLint,BL is inhibited by 90%, hepatic E217G exposure is 2.3-fold (point B). However, when both CLint,bile and CLint,BL are inhibited by 90%, the hepatic E217G exposure increases 10-fold (point C).
Discussion
Understanding hepatic E217G disposition is important because E217G plays a role in ICP and susceptibility could change if transporter expression is altered by disease or drug interactions. Therefore, this study characterized E217G disposition in SCHH using the uptake and efflux protocol and mechanistic modeling previously established (Pfeifer et al., 2013) to estimate basolateral uptake and efflux clearance values and biliary excretion clearance. These values were used to study the relative contribution of the different pathways and simulate effects of cholestasis-mediated transporter alterations or drug-drug interactions on hepatic E217G disposition.
The biliary excretion of E217G by Mrp2 is well documented (Huang et al., 2000; Morikawa et al., 2000). The influence of biliary excretion on E217G disposition in SCHH is indicated by the BEI, which ranged from 40% to 52% in the three hepatocyte lots. This BEI range for E217G agrees with previously published values of 32%–43% (Bi et al., 2006; Lee et al., 2010). The mean E217G CLint,bile estimate determined here [0.11 (ml/min)/g liver] was similar to TCA CLint,bile [0.14 (ml/min)/g liver] determined previously (Yang et al., 2015). Whereas TCA is predominantly excreted into bile in SCHH (Yang et al., 2015), this is not the case for E217G. The CLint,BL of TCA is 3.3-fold lower than CLint,bile (Yang et al., 2015). In contrast, based on our results, the CLint,BL of E217G is 1.6-fold higher than CLint,bile (Table 2). Thus, basolateral efflux is an important route of E217G elimination in SCHH. It should be noted that there is inter-donor variability in the ratio of CLint,BL to CLint,bile; the ratio is 0.9, 2.0, and 2.9 in hepatocyte Lots 1, 2, and 3, respectively (Table 2). This variability is expected based on large individual differences in protein levels of hepatic efflux transporters. For example, MRP2 and MRP3 protein expression showed 6.6- and 3.7-fold individual differences, respectively, in human liver tissue from more than 50 donors (Deo et al., 2012; Wang et al., 2015).
At the basolateral membranes of hepatocytes, MRP3 and MRP4 transport substrates from the cells to the sinusoidal blood (Chandra and Brouwer, 2004). Although our data cannot identify the transporters responsible for the basolateral efflux of E217G, it is likely to be mediated by MRP3 and/or MRP4, because E217G is a well-established substrate for these efflux transporters (Zeng et al., 2000; Chen et al., 2001). MRP3 and MRP4 have comparable Km values for E217G (Zeng et al., 2000; Chen et al., 2001; Zamek-Gliszczynski et al., 2006). However, in human liver tissue, the absolute protein level of MRP3 is at least 40-fold higher than MRP4 (Vildhede et al., 2015). Thus, MRP3 is assumed to be the primary transporter mediating the hepatic basolateral efflux of E217G in healthy subjects. It should be noted that E217G is not a substrate of organic solute transporter α/β, which also is expressed on the basolateral membranes of hepatocytes (Malinen et al., 2018).
The contribution of basolateral efflux to E217G elimination from hepatocytes could be amplified in situations where basolateral efflux transporter levels are increased. Basolateral transporters can be induced in several liver diseases, including cholestasis (Thakkar et al., 2017). For example, MRP3 protein levels increased 4.6-fold in patients with obstructive cholestasis caused by gall stones (Chai et al., 2012) and increased 2.1- to 2.6-fold in patients with stage III/IV primary biliary cirrhosis (Zollner et al., 2003). A similar increase compared with control subjects was reported for MRP4 in obstructive cholestasis (3.1-fold) and primary biliary cirrhosis (3.2- to 3.5-fold) (Zollner et al., 2007; Chai et al., 2011). MRP3 and MRP4 protein levels also are increased in nonalcoholic steatohepatitis (Hardwick et al., 2011). In addition, Mrp3/MRP3 is upregulated in Mrp2-deficient TR− rats and in patients with Dubin-Johnson syndrome (König et al., 1999; Johnson et al., 2006). In contrast, MRP2 levels do not appear to be changed in cholestasis in humans, but some studies point to a mislocalization of MRP2 in cholestatic disease (Zollner et al., 2001, 2003; Kojima et al., 2003; Chai et al., 2012).
A sensitivity analysis was performed to evaluate the effects of changes in efflux protein levels on hepatic E217G disposition. The analysis revealed that hepatic E217G was sensitive to CLint,BL; a fourfold increase in CLint,BL decreased hepatic E217G by 64% (Fig. 4A). Biliary E217G also was examined because E217G in bile may play a role in the development of cholestasis through trans-inhibition of BSEP (Stieger et al., 2000). Biliary E217G was influenced by both basolateral and biliary transporters (Fig. 4B). Sensitivity analysis of CLint,bile revealed that a decrease in CLint, bile (e.g., MRP2 downregulation) effectively decreased the amount of E217G in bile (Fig. 4B). However, the effects on E217G accumulation in hepatocytes were smaller (Fig. 4A). Importantly, decreased CLint,bile caused only a minimal increase in cellular E217G amounts when CLint,BL was induced fourfold (Fig. 4A). These results suggest that induction of basolateral efflux transporters (e.g., MRP3 and MRP4) in cholestasis could lower the amount of E217G in hepatocytes, even when biliary clearance is impaired, and may help relieve cholestasis. Pregnant women with defects in these compensatory mechanisms could be predisposed to developing ICP.
The inhibition of MRP3 and MRP4 is a risk factor for drug-induced cholestasis (Morgan et al., 2013; Köck et al., 2014; Chan and Benet, 2018). In addition, MRP2 inhibition alongside possible mislocalization in cholestasis can disturb hepatocellular homeostasis of MRP2 substrates. As expected, our simulations revealed that inhibition of either CLint,BL alone (i.e., MRP3 and MRP4 transport) or inhibition of CLint,bile alone (i.e., MRP2 transport) has a minimal effect on hepatic E217G exposure (Fig. 5). This is consistent with the concept of fractional excretion (Zamek-Gliszczynski et al., 2009). Based on the calculated clearance estimates in SCHH, 62% of E217G is excreted via the basolateral pathway. Therefore, even a complete knockout of this predominant excretion pathway would only increase E217G in hepatocytes by a maximum of 2.6-fold, assuming that no changes in the compensatory biliary pathway occurred. However, simulations indicated that hepatic E217G was markedly increased with simultaneous inhibition of both pathways (Fig. 5). The simultaneous inhibition of several MRP transporters is a likely event due to overlap in the inhibitor specificity of MRPs (Morgan et al., 2013), and this may predispose patients to E217G-mediated cholestasis. For example, medications that have been associated with drug-induced liver injury and cholestasis (e.g., repaglinide, cyclosporine A, everolimus) exhibit overlapping inhibition of MRP2, MRP3, and MRP4 (Morgan et al., 2013).
The hepatic clearance estimates of E217G were determined assuming linear processes. No saturation of these pathways is expected with the 0.3 µM E217G dose used in this study. The apparent Km for the uptake clearance of E217G in plated human hepatocytes is reported to be 7.7 µM (Liao et al., 2019), and the Km values of E217G for MRP3 and MRP4 are in the range of 20–40 µM determined in vesicle studies (Zeng et al., 2000; Chen et al., 2001; Zamek-Gliszczynski et al., 2006). E217G shows cooperative binding and sigmoidal kinetics of MRP2-mediated transport in membrane vesicles with Km/K0.5 values of >100 µM (Herédi-Szabó et al., 2009). The highest hepatocellular concentrations of E217G reached in this study were <1.5 µM. Therefore, we believe that the assumption of linearity is reasonable. Another simplification of the model was that passive permeability was not modeled as a separate parameter. However, passive permeability has been shown previously to play a minor role in E217G uptake in hepatocytes, with active processes contributing to 89.3% ± 8.1% of total uptake (Liao et al., 2019). The biliary excretion of E217G was decreased by more than 75% in the presence of the MRP2 inhibitor sulindac or its metabolite (Lee et al., 2010), suggesting that passive permeability is also minor in E217G efflux from hepatocytes.
SCHH continue to be a standard method for assessing hepatic disposition in vitro, particularly regarding transporter-mediated efflux (Brouwer et al., 2013). As bile networks re-form over days in culture, the levels of efflux transporters increase and exhibit appropriate functionality (Swift et al., 2010). An important consideration with this study is that levels of individual transporters in SCHH may differ from those in liver tissue, which could limit the ability to accurately translate data from in vitro to in vivo. Recently, the absolute protein abundance of MRP2 and MRP3, the primary transporters involved in hepatic E217G efflux, was reported to be approximately fourfold higher, on average, in SCHH than in liver tissue from corresponding donors (Kumar et al., 2019). However, the ratio of absolute levels of MRP2 to MRP3 was similar in SCHH and liver tissue (Kumar et al., 2019). Although protein abundance was not determined in the current study, the available literature suggests that the relative contribution of the efflux pathways of interest are similar in SCHH and in liver tissue.
In summary, this study revealed that basolateral efflux in addition to biliary excretion is a significant route of E217G elimination in SCHH. The importance of this elimination pathway is further emphasized when basolateral efflux transporter levels (e.g., MRP3 and MRP4) are increased, which may occur in conditions such as cholestasis. The simulations presented here highlight how hepatocytes can effectively cope with increased E217G through the regulation of both basolateral and biliary transporters.
Acknowledgments
The authors acknowledge Certara for providing Phoenix software to the UNC Eshelman School of Pharmacy, Division of Pharmacotherapy and Experimental Therapeutics, as part of the Pharsight Academic Center of Excellence Program.
Authorship Contributions
Participated in research design: Ito, Brouwer.
Conducted experiments: Ito, Sjöstedt.
Performed data analysis: Ito, Sjöstedt, Brouwer.
Wrote or contributed to the writing of the manuscript: Sjöstedt, Ito, Brouwer.
Footnotes
- Received July 31, 2019.
- Accepted November 12, 2019.
↵1 K.I. and N.S. contributed equally to this work.
This work was supported by the National Institute of General Medical Sciences of the National Institutes of Health [Award Number R35 GM122576] (K.L.R.B.) and by Teijin Pharma Limited. N.S. was supported by the Sigrid Jusélius Foundation.
This work was presented, in part, at the AAPS 2018 PharmSci 360 (Washington, DC, November 4-7, 2018).
K.L.R.B. is a co-inventor of the sandwich-cultured hepatocyte technology for quantification of biliary excretion (B-CLEAR©) and related technologies, which have been licensed exclusively to Qualyst Transporter Solutions, recently acquired by BioIVT. B-CLEAR© is covered by U.S. patent 6,780,580 and other U.S. and international patents both issued and pending.
Abbreviations
- BEI
- biliary excretion index
- BSEP
- bile salt export pump
- CLint,bile
- intrinsic biliary excretion clearance
- CLint,BL
- intrinsic basolateral efflux clearance
- CLuptake
- uptake clearance
- E217G
- estradiol-17β-glucuronide
- HBSS
- Hanks’ balanced salt solution
- ICP
- intrahepatic cholestasis of pregnancy
- Kflux
- rate constant for flux from bile into the medium
- MRP
- multidrug resistance-associated protein
- OATP
- organic anion transporting polypeptide
- SCHH
- sandwich-cultured human hepatocytes
- TCA
- taurocholate
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}