


Abstract
Cytochrome P450 (P450) and uridine 5′-diphospho-glucuronosyltransferase (UGT) enzymes mediate a major proportion of phase I and phase II metabolism of xenobiotics. In vitro-in vivo extrapolation (IVIVE) of hepatic clearance in conjunction with physiologically-based pharmacokinetics (PBPK) has become common practice in drug development. However, prediction of xenobiotic kinetics in virtual populations requires knowledge of both enzyme abundances and the extent to which these correlate. A multiplexed quantification concatemer (QconCAT) strategy was used in this study to quantify the expression of several P450 and UGT enzymes simultaneously and to establish correlations between various enzyme abundances in 24 individual liver samples (ages 27–66, 14 male). Abundances were comparable to previously reported values, including CYP2C9 (40.0 ± 26.0 pmol mg−1), CYP2D6 (11.9 ± 13.2 pmol mg−1), CYP3A4 (68.1 ± 52.3 pmol mg−1), UGT1A1 (33.6 ± 34.0 pmol mg−1), and UGT2B7 (82.9 ± 36.1 pmol mg−1), expressed as mean ± S.D. Previous reports of correlations in expression of various P450 (CYP3A4/CYP3A5*1/*3, CYP2C8/CYP2C9, and CYP3A4/CYP2B6) were confirmed. New correlations were demonstrated between UGTs [including UGT1A6/UGT1A9 (rs = 0.82, P < 0.0001) and UGT2B4/UGT2B15 (rs = 0.71, P < 0.0001)]. Expression of some P450 and UGT enzymes were shown to be correlated [including CYP1A2/UGT2B4 (rs = 0.67, P = 0.0002)]. The expression of CYP3A5 in individuals with *1/*3 genotype (n = 11) was higher than those with *3/*3 genotype (n = 10) (P < 0.0001). No significant effect of gender or history of smoking or alcohol use on enzyme expression was observed; however, expression of several enzymes declined with age. The correlation matrix produced for the first time by this study can be used to generate more realistic virtual populations with respect to abundance of various enzymes.
Introduction
In vitro-in vivo extrapolation (IVIVE) is an integral part of physiologically-based pharmacokinetics (PBPK) in modern drug development. However, IVIVE requires scaling factors to relate drug-related values obtained from in vitro experiments to human biology and clinical observations. In particular, the absolute abundance of hepatic drug-metabolizing enzymes is required to extrapolate metabolism rates determined from recombinantly expressed enzymes to in vivo drug clearance (Barter et al., 2007). Virtual populations created using Monte-Carlo simulations can facilitate understanding the interindividual variability of pharmacokinetic parameters (Rostami-Hodjegan and Tucker, 2007), but traditionally, and mainly owing to lack of data, the values assigned to each enzyme in a given virtual subject are independent of each other, with few exceptions (CYP3A4 and 3A5) (Barter et al., 2010). Many cytochrome P450 (P450) enzymes share expression-regulation mechanisms, and therefore correlations in expression can be expected (Jover et al., 2009). For example, the aryl-hydrocarbon nuclear receptor (AhR) (Hankinson, 1995), hepatocyte nuclear factor 4 (HNF-4) (Venepally et al., 1992), constitutive androstane receptor (CAR) (Baes et al., 1994), and pregnane X receptor (PXR) (Lehmann et al., 1998; Burk et al., 2004) are reported to be involved in the regulation of CYP1A, CYP2A/2C/2D, CYP2B, and CYP3A subfamilies, respectively.
Western blotting is an established method to measure the abundance of cytochrome P450 (Laemmli, 1970; Towbin et al., 1979), but has many disadvantages. Most important in the case of the highly homologous enzymes within the cytochrome P450 and 5′-diphospho-glucuronosyltransferase (UGT) families (Fallon et al., 2008) is the dependence on the quality of standards and specificity of antibodies. Nevertheless, immunoquantification methods have been used extensively to measure the abundance of cytochrome P450 enzymes (Shimada et al., 1994; Yamazaki et al., 1997; Tateishi et al., 1999; Lin et al., 2002).
Liquid chromatography in conjunction with mass spectrometry (LC-MS)–based quantitative assays are an attractive alternative to immunoassays. Mass spectrometry is an extraordinarily sensitive and selective technique, routinely detecting femtomoles of analytes differing in mass by fractions of a Dalton. The LC and tandem mass spectrometry (MS/MS) steps introduce further resolution based on hydrophobicity and (in the case of peptides) on sequence, respectively. An isotopically-labeled standard corresponding to each analyte under study is introduced to carry out internal standard quantification of analyte proteins and to account for intersample variation and matrix effects. Several LC-MS/MS methods have been recently used to measure the abundance of cytochrome P450 (Seibert et al., 2009; Kawakami et al., 2011; Ohtsuki et al., 2012) and UGT (Fallon et al., 2008, 2013; Sato et al., 2012; Sridar et al., 2013) enzymes.
The quantification concatemer (QconCAT) technique was developed to quantify several proteins simultaneously in each sample and has not previously been applied to drug-metabolizing enzymes and transporters. The method involves an artificial protein comprising concatenated proteotypic signature peptides for a targeted set of proteins. The QconCAT is expressed in Escherichia coli in an isotopically enriched medium (Beynon et al., 2005), then spiked into a protein mixture, where, on proteolytic digestion, it yields isotopically labeled standard peptides for each of the targeted proteins, in equimolar concentration (Simpson and Beynon, 2012). The synthesis and expression of a suitable QconCAT involves selection of suitable well-behaved proteotypic peptides and subsequent expression of proteins and has a significant failure rate (about 20%); strategies for expression of recalcitrant QconCATs include reordering (reshuffling) the peptides or linking the QconCAT to a fusion partner (Russell et al., 2013). However, once the hurdles of optimizing the QconCAT are resolved the full range of cytochrome P450 and UGT enzymes (or transporters) can be measured simultaneously in the same samples. QconCAT presents some advantages over the commonly used synthetic AQUA peptides, including the possibility of equimolar multiplexed quantification and the lower cost of a sustainable supply of standard peptides (Beynon et al., 2005).
Simultaneous measurement of the enzymes and transporters facilitates creation of more realistic virtual populations as part of physiologically-based pharmacokinetic–IVIVE models because the common effects exerted by genetic and environmental factors are not always independent of one another. The expression and activity of drug-metabolizing enzymes and transporters can be correlated and correlations affecting drug absorption, distribution, metabolism, and excretion as well as absolute values are used to inform in silico models. To elucidate the correlations between drug-metabolizing enzymes, the abundances of a wide range of these enzymes have to be assessed in the same set of samples.
This study developed a selected reaction monitoring (SRM)–based quantitative assay, which was validated and used to quantify multiple cytochrome P450 (13) and UGT (8) enzymes in 24 human liver microsomal (HLM) samples. Correlations of expression between the quantified enzymes were investigated and the effects of certain genetic and environmental factors on the measured abundances of these enzymes were explored.
Materials and Methods
Materials and Chemicals
All materials and chemicals were purchased from Sigma-Aldrich (Poole, UK) unless otherwise indicated. Lysyl endopeptidase (Lys-C) was purchased from Wako (Osaka, Japan). The light version of the modified Glu-fibrinopeptide was purchased from Cambridge Peptides (Cambridge, UK). The QconCAT used in this study, metabolism enzymes quantification concatemer (MetCAT), was produced in-house as previously described (Russell et al., 2013).
Human Liver Microsomal Samples
Human liver microsomes included one pooled sample of human liver microsomes (pool of 50 livers; BD Gentest/BD Biosciences, San Jose, CA) and 23 individual HLM samples from Caucasian subjects and one from an African-American subject (BD Gentest/BD Biosciences), supplied with CYP3A4 and CYP3A5 abundance determined by either Western blotting (pooled HLM sample) or enzyme-linked immunosorbent assay (ELISA) (individual HLM samples) and reported previously (Barter et al., 2010). Table 1 shows demographic details of the 24 individual sample donors (24 individual HLM samples were randomly selected from a set of 52 samples, 24 being the largest number of samples that could realistically be processed in a single experiment).
Demographic details of the 24 individual liver donors
The MetCAT Standard
The MetCAT standard has previously been described (Russell et al., 2013). The tryptic peptides making up the QconCAT were two each from 15 cytochrome P450 and 10 UGT enzymes (CYP1A2, CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2D6, CYP2E1, CYP3A4, CYP3A5, CYP3A7, CYP3A43, CYP2J2, CYC4F2 and UGT1A1, UGT1A3, UGT1A4, UGT1A6, UGT1A9, UGT2B4, UGT2B7, UGT2B10, UGT2B11, UGT2B15). Peptides unique to subfamilies CYP1A, CYP2A, CYP2B, CYP3A, and UGT1A were also included to enable aggregate quantification of each subfamily. Finally, a Glu-fibrinopeptide B analog (GVNDNEEGFFSAR) was included to enable quantification of the MetCAT itself (Eyers et al., 2008), hereafter referred to as NNOP (nonnaturally occurring peptide). All the required peptides are concatenated together in the MetCAT protein and are released on proteolytic digestion (Supplemental Table 1).
Sample and Standard Preparation
Microsomal protein concentration was determined in triplicate with Bradford assay (Bradford, 1976) following the manufacturer’s instructions. Microsomal protein was solubilized as previously described (Achour et al., 2011). Briefly, 5 μl of microsomal sample was diluted in 20 μl double-distilled H2O then 25 μl of SDS loading buffer was added (final concentrations: Tris-HCl 100 mM, pH 6.8; SDS 5%; glycerol 15%; β-mercaptoethanol 7.5%; EDTA 0.5 mM; and bromophenol blue 0.002%). The mixture incubated at room temperature for 1 hour, then heated for 10 minutes at 95°C. Both samples and standard were resolved by a 10% w/v SDS-PAGE gel on a mini-Protean 3 system (Bio-Rad, Hercules, CA). Bands were visualized by Coomassie brilliant blue staining (0.25% w/v Coomassie brilliant blue R-250, 45% v/v methanol, 10% v/v acetic acid, 45% v/v H2O).
In-Gel Digestion
The microsomal cytochrome P450 and UGT region (45–65 kDa) and the MetCAT band were excised and cut into pieces of approximately 2 × 2 mm, destained with two washes of 150 μl 50% acetonitrile in 25 mM ammonium bicarbonate, reduced with 200 μl of 10 mM dithiothreitol in 50 mM ammonium bicarbonate at 50°C for 30 minutes, alkylated with 100 μl of 55 mM iodoacetamide in 50 mM ammonium bicarbonate in the dark for 1 hour, flushed with two washes of 200 μl of 50 mM ammonium bicarbonate, dehydrated once with 100 μl acetonitrile, and air dried for 10 minutes.
Protein was digested in 25 mM ammonium bicarbonate buffer, firstly with lysyl endopeptidase (20 μl, 0.01 μg μl−1); the volume was adjusted to 50 μl and samples incubated overnight at 30°C. A second digestion was then carried out, using trypsin (20 μl, 0.01 μg μl−1); the volume was adjusted to 80 μl and the mixture incubated overnight at 37°C. Peptides were recovered by retaining the aqueous solution from the digest, then washing the gel pieces twice for 15 minutes with 100 μl 5% acetonitrile and then 100 μl 50% acetonitrile, and adding these washings to the retained aqueous solution. Sample volume was reduced to less than 50 μl using a vacuum concentrator (Eppendorf, Hamburg, Germany) and then adjusted to 50 μl with 2% acetonitrile, 0.1% formic acid in water, and stored at –20°C.
Preparation of the Sample-Standard-NNOP Assay Mix for the SRM Assay
Mixtures of sample, standard, and NNOP were prepared as follows: 15 μl of 1 in 2 diluted sample (in 3% acetonitrile, 0.1% formic acid), 3 μl of 1 in 10 diluted MetCAT, and 1.5 μl of NNOP (17.15 fmol μl−1 diluted 1:200,000 from stock; 10.3 mg in 2 ml at purity of 95.94%).
SRM of QconCAT-Sample Mixtures
Samples were analyzed by LC-MS/MS using a nanoACQUITY nano-HPLC system (Waters, UK) coupled to a TSQ Vantage triple quadrupole mass spectrometer (ThermoScientific, Pittsburgh, PA). Selected reaction monitoring assays were designed and managed using Skyline version 1.4.0.4222 (MacCoss Laboratory Software, Department of Genome Sciences, University of Washington, Seattle, WA) (MacLean et al., 2010) and data acquired using Xcalibur version 2.0.6 Service Pack 1 (Thermo Fisher). Samples (3 μl, 30 fmol analyte, 3% acetonitrile, 0.1% formic acid) were injected using the partial loop mode onto a trapping column (Symmetry C18, 180 μm × 20 mm; Waters), at flow rate of 5 μl min−1 for 3 minutes. The trap was then brought in line with the analytical column (1.8 μm HSS T3, 75 μm × 150 mm; Waters) maintained at 35°C. Peptides were eluted at a 0.3 μl min−1 flow rate and a gradient of 3–50% acetonitrile over 40 minutes followed by a ramp to 95% acetonitrile for 3 minutes then a return to starting conditions for 14 minutes to re-equilibrate the system.
Statistical Analysis of the Measured Microsomal Enzyme Abundance Data
Calculation of Enzyme Abundance Values
The abundance of each enzyme was calculated with the following equation: (1)where AProtein is the estimate of protein abundance in starting microsomal material, based on the surrogate peptide abundance estimate, measured in units of pmol mg−1 microsomal protein. [NNOP] is the concentration of the light NNOP in the assay mix in fmol μl−1 and RH.NNOP/L.NNOP the ratio of heavy, MetCAT-derived NNOP to light (standard of known concentration) NNOP; together these terms quantify the MetCAT equimolar concentration. RL.peptide/H.peptide is the ratio of light (sample-derived) analytical peptide to heavy (MetCAT-derived) peptide, combined with previous terms to give peptide concentration in the assay mix in fmol μl−1. FSample is the dilution factor of the microsomal sample in the mixture (7.5 μl in 19.5 μl for all individual HLM samples) and the term containing it converts the peptide concentration in the assay mix to the concentration in the digest. VDigest is the digest volume in μl, converting the peptide concentration into an absolute abundance in fmol. PCMicrosomal is the microsomal protein content corresponding to the amount loaded on the gel prior to digestion expressed in μg, the term containing it relates abundance in the digest to abundance in microsomal protein in pmol mg−1. The ratios RH.NNOP/L.NNOP and RL.peptide/H.peptide were corrected for isotope incorporation efficiency prior to use in this equation. In the case where the pair of peptides representing the same protein returned different quantitative ratios, the peptide that provided the higher abundance was used in line with published QconCAT protocols (Carroll et al., 2011).
(1)where AProtein is the estimate of protein abundance in starting microsomal material, based on the surrogate peptide abundance estimate, measured in units of pmol mg−1 microsomal protein. [NNOP] is the concentration of the light NNOP in the assay mix in fmol μl−1 and RH.NNOP/L.NNOP the ratio of heavy, MetCAT-derived NNOP to light (standard of known concentration) NNOP; together these terms quantify the MetCAT equimolar concentration. RL.peptide/H.peptide is the ratio of light (sample-derived) analytical peptide to heavy (MetCAT-derived) peptide, combined with previous terms to give peptide concentration in the assay mix in fmol μl−1. FSample is the dilution factor of the microsomal sample in the mixture (7.5 μl in 19.5 μl for all individual HLM samples) and the term containing it converts the peptide concentration in the assay mix to the concentration in the digest. VDigest is the digest volume in μl, converting the peptide concentration into an absolute abundance in fmol. PCMicrosomal is the microsomal protein content corresponding to the amount loaded on the gel prior to digestion expressed in μg, the term containing it relates abundance in the digest to abundance in microsomal protein in pmol mg−1. The ratios RH.NNOP/L.NNOP and RL.peptide/H.peptide were corrected for isotope incorporation efficiency prior to use in this equation. In the case where the pair of peptides representing the same protein returned different quantitative ratios, the peptide that provided the higher abundance was used in line with published QconCAT protocols (Carroll et al., 2011).
Analysis of Variance of the Enzyme Abundances in the Pooled HLM Sample
The precision of the assay was investigated with one-way analysis of variance (ANOVA), which enabled variability in estimate of enzyme abundance to be partitioned between sample preparation and the SRM assay itself. A single pooled HLM sample was prepared in triplicate and the cytochrome P450 enzymes in each replicate quantified by the SRM assay also in triplicate. The ANOVA was performed using GraphPad InStat software version 3.05 (GraphPad Software, San Diego, CA). Here and throughout, P values ≤ 0.05 are considered significant.
Establishing Linearity of the Quantification Assay
Linear regression analysis was carried out to test linearity between the concentration of the light NNOP and its light-to-heavy peak intensity ratio and between the concentration of sample peptides and the dilution factor used to prepare the sample.
Detected peptides with quantitative ratios of light-to-heavy peak intensity ranging from 0.1 to 10 were selected for the linearity test (sample versus QconCAT). Measured concentration was plotted against the dilution factor of the sample; 1 in 1, 1 in 2, 1 in 4, 1 in 8, 1 in 16, and 1 in 32 (plotted as fractions: 1, 1/2, 1/4, 1/8, 1/16, and 1/32). All volumes were kept the same to limit errors that can be introduced into the assay (10 μl of sample, 2 μl of 1 in 10 heavy labeled MetCAT, and 1 μl light NNOP standard peptide).
Analysis of Enzyme Abundances in the Individual Microsomal Samples
Means, standard deviations, and coefficients of variation of abundances were calculated. Normality of distribution of the abundance measurements of each enzyme were tested according to the method of Kolmogorov and Smirnov.
Correlation Analysis of the Measured Enzyme Abundances
In this case, the normality tests were mostly negative (see Results), and therefore the nonparametric Spearman rank test (Armitage et al., 2001) was used to assess correlations of the expression levels of the enzymes. Correlations were also tested for abundances and activities of the enzymes CYP1A2, CYP2B6, CYP2C9, CYP2D6, and CYP3A4 (data for activities provided by Pfizer, New York, NY). Activities had been measured at single concentrations of the enzymes according to previously published methods (Walsky and Obach, 2004) using the following probes: phenacetin (CYP1A2), mephenytoin (CYP2B6), diclofenac (CYP2C9), bufuralol (CYP2D6), and testosterone (CYP3A), and the data were used in several previous publications (e.g., Walsky et al., 2012). Calculations of the correlation coefficients, rs, and the probabilities, P, were carried out using GraphPad InStat software version 3.05. Graphs were generated using GraphPad Prism software version 5.04 (GraphPad Software).
Analysis of the Effects of Genotype, Age, Gender, Smoking, and Alcohol Use on Measured Abundance of Enzymes
Genotypes were available for CYP2B6, CYP2C9, CYP2D6, and CYP3A5. The abundance data of these four enzymes were stratified according to genotype. The one-way nonparametric Kruskal-Wallis ANOVA rank test was applied where there were more than two genotypes; otherwise, the unpaired Mann-Whitney rank test was used. Post-hoc statistical analysis for the Kruskal-Wallis ANOVA tests was carried out using multiple pairwise Mann-Whitney tests with Bonferroni corrected P values. Nonparametric Spearman rank correlation analysis was performed to identify relationships between cytochrome P450 and UGT enzyme abundances and the age of donors. Additionally, effects of gender, smoking, and alcohol consumption on abundances of enzymes were assessed using Mann-Whitney rank U-test.
The supplier provided demographic and clinical details including age, gender, medical history, history of smoking, and alcohol consumption, but not medication history of donors. One subject was stated to be a smoker but had stopped smoking 11 years before donation. It is well known that induction of CYP1A2 by smoking wears off after five half-lives (Faber and Fuhr, 2004). Therefore, this subject was considered to be a nonsmoker for the purpose of our analysis.
Results
A QconCAT-based assay was successfully developed to quantify several P450 and UGT enzymes relevant to the metabolism of xenobiotics in one pooled HLM sample and 24 HLM samples.
Accuracy, Precision, and Linearity of the SRM Quantification Assay
Linearity of the assay was confirmed by applying the assay to quantification of the P450 enzymes in a series of 2-fold dilutions of peptides derived from a single digest of the pooled HLM sample. The set of measured concentrations demonstrated linearity over a range of 1- to 32-fold dilution (illustrated for CYP3A4 and CYP3A5 in Fig. 1A).

Linearity, precision, and accuracy of measurement of CYP3A4 and CYP3A5 in the pooled HLM sample using the developed assay (A). The concentration of the two peptides representing CYP3A4 and CYP3A5 showed strong linear correlation over a wide dilution range. The undiluted sample was used to test precision and showed limited variability between nine individual measurements represented by the error bars (standard deviations). Good agreement seen between measurements of the two enzymes using LC-MS and Western blotting for CYP3A4 measurement (B). In B: dashed line, line of identity; solid line, line of regression.
For the purpose of accuracy testing, Western blot–derived estimates of CYP3A4 and CYP3A5 were provided by the supplier of the pooled sample and ELISA-derived estimates of CYP3A4 in five individual HLM samples (Barter et al., 2010) were also available. All the CYP3A4 estimates were compared with values obtained using the LC-MS assay (Fig. 1B). Linear regression showed very good correlation between the two sets of measurements.
Precision of the assay was assessed with an ANOVA analysis in which the pooled sample was prepared in triplicate and each preparation assayed by LC-MS in triplicate. This revealed the extent of random error in the assay and partitioned the source of that error between the sample preparation and the LC-MS analysis. The ANOVA-based F values (Table 2) were statistically significant and greater than one, indicating that the greater source of random error is sample preparation rather than LC-MS analysis. However, the fold differences between minimum and maximum mean expression estimates for each set of replicate sample preparations was still low at 1.07–1.35, giving some confidence in the robustness of the assay. Reproducibility was confirmed by the coefficient of variation of nine measurements of each of the 14 peptides in the sample; this coefficient of variation did not exceed 18%.
Summary of the ANOVA analyses on measured concentrations of 14 cytochrome P450 Q-peptides
The pooled sample was prepared three times and each prepared sample set was run on LC–triple quadrupole mass spectrometer (QqQ) three times. P values were computed to assess the overall difference between measurements, and the F test was used to assess sources of variation. Fold differences and coefficients of variation of measurements of each peptide are shown.
Enzyme Abundances in Individual HLM Samples
The SRM assay was applied to estimate cytochrome P450 (CYP1A2, 2A6, 2B6, 2C8, 2C9, 2C18, 2D6, 2J2, 3A4, 3A5, 3A7, 3A43, and 4F2) and UGT (UGT1A1, 1A3, 1A4, 1A6, 1A9, 2B4, 2B7, and 2B15) enzyme expression in 24 individual samples. A small number of peptides were not detected (in heavy and light versions) in all samples and some returned ratios outside the quantitative range.
The proteins CYP2C19, CYP2E1, UGT1A10, and UGT1A11 produced elution profiles of inadequate quality in these experiments and therefore were not considered for measurement at this time. CYP2A6, CYP2C9, CYP2D6, CYP2J2, CYP3A7, CYP4F2, UGT1A3, and UGT1A9 were quantified using a single peptide. There are always unforeseen circumstances affecting LC-MS quantification, and redundancy is a feature of the QconCAT strategy. CYP1A2, CYP2B6, CYP2C8, CYP2C18, CYP3A4, CYP3A43, UGT1A1, UGT1A4, UGT1A6, UGT2B4, and UGT2B15 were, however, each quantified by two peptides in the QconCAT sequence (giving six transitions) and it would be expected that both peptides should give the same result. Although technical replicates for these measurements were in good agreement, in the cases of CYP2B6 and UGT1A6, the two peptides did not produce equivalent results. In these cases, the higher value of the quantification was considered to be the more accurate.
Tables 3 and 4 show summaries of cytochrome P450 and UGT abundance data, respectively. Pie charts of P450 and UGT enzyme abundances in the liver are shown in Supplemental Figs. 1 and 2. Large interindividual variation is observed in the measured abundances of the enzymes, which are poorly represented by the mean values (Fig. 2).
The arithmetic means, the standard deviations (S.D.), percent coefficients of variation (CV), and the ranges of the measured abundances of 13 cytochrome P450 enzymes
The arithmetic means, the standard deviations (S.D.), percent coefficients of variation (CV), and the ranges of the measured abundances of eight UGT enzymes
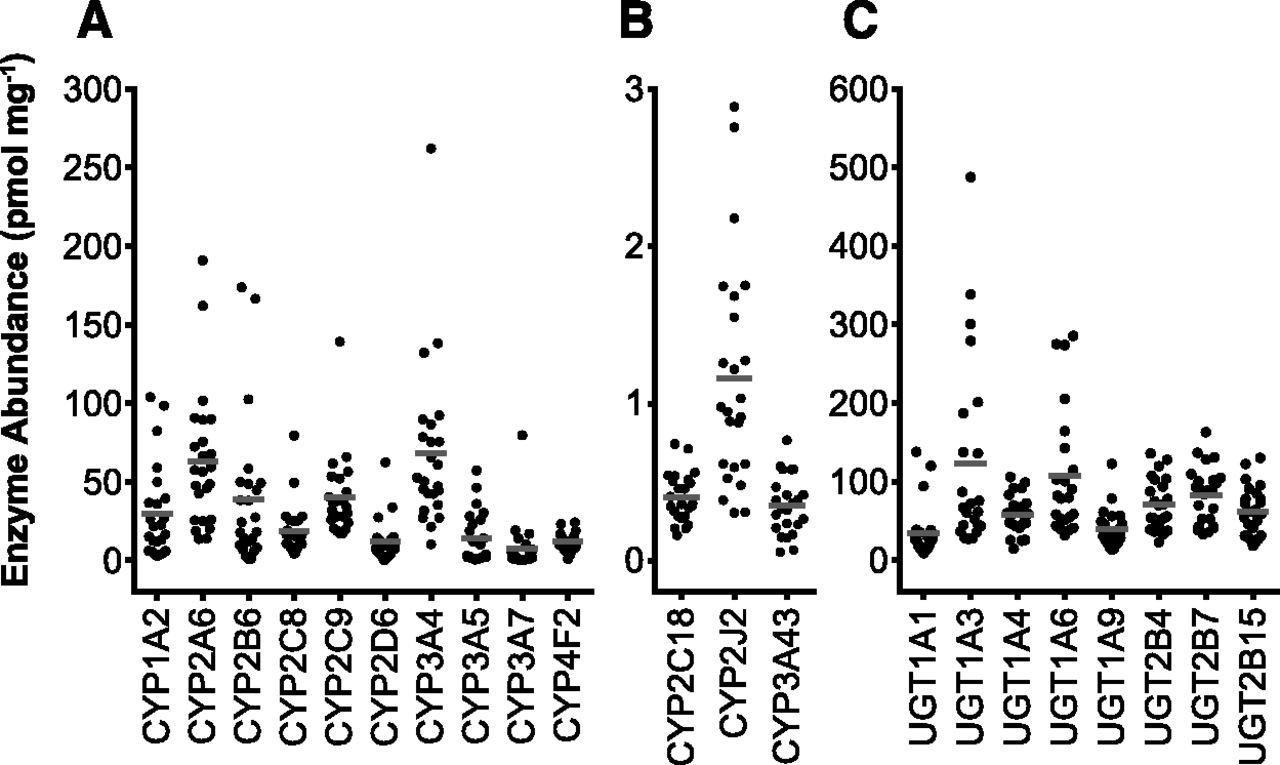
A scatter plot of the measured abundance values of P450 (A and B) and UGT (C) enzymes. The number of samples is 24 for each enzyme except CYP2C9, CYP3A5, CYP3A7, CYP3A43, UGT1A3, UGT1A4, and UGT1A6 (n = 23). Lines indicate population means of the sets of data.
Correlation Analysis of Enzyme Expression
Pairwise correlation in enzyme expressions was tested with the nonparametric Spearman rank test (Table 5, example correlation plots in Fig. 3), which is tolerant both to deviation from normality and to outlier data points (Armitage et al., 2001). This test was chosen because initial tests for normality in expression distribution of enzyme abundance data, using the Kolmogorov and Smirnov test, indicated statistically significant deviation from normality for all but eight of the enzymes (CYP2A6, CYP2C18, CYP2J2, CYP3A43, CYP4F2, UGT1A4, UGT2B7, and UGT2B15), and this may have been due to low sensitivity of the test with this sample size.
Summary matrix of correlation analysis of cytochrome P450 and UGT abundance data
Statistically nonsignificant correlations are not shown (—). n = 24, except for correlations of CYP2C9, 3A5, 3A7, 3A43, UGT1A3, 1A4, and 1A6 (n = 23).

Correlation plots for CYP3A5 and CYP3A4 (A), CYP2B6 and CYP3A4 (B), CYP2C8 and CYP2C9 (C), and UGT1A9 and UGT1A6 (D). Correlation between CYP3A5*1/*3 (A, open circles) and CYP3A4 levels is statistically significant and strong (rs = 0.92, P < 0.0001, n = 12) and correlation is weak when CYP3A5*3/*3 variant abundance data (A, closed circles) are included (rs = 0.36, P = 0.09, n = 23). Strong correlations exist between the pairs: CYP2B6/CYP3A4 (rs = 0.63, P = 0.0004, n = 24), CYP2C8/CYP2C9 (rs = 0.56, P = 0.003, n = 23), and UGT1A6/UGT1A9 (rs = 0.82, P < 0.0001, n = 23).
Relationship between Abundance of Enzymes and Genotype
Figure 4 shows the abundance of P450 enzymes stratified by genotype. There was a significant difference between expression levels of CYP3A5*1/*3 and CYP3A5*3/*3 (Mann-Whitney U-test; P < 0.0001). However, there were no statistically significant differences between expression levels of the different genotypes of CYP2B6 and CYP2C9 (nonparametric ANOVA; P = 0.38 and P = 0.58, respectively). Differences in CYP2D6 expression were of borderline significance; however, post-hoc analysis showed this difference was not significant.
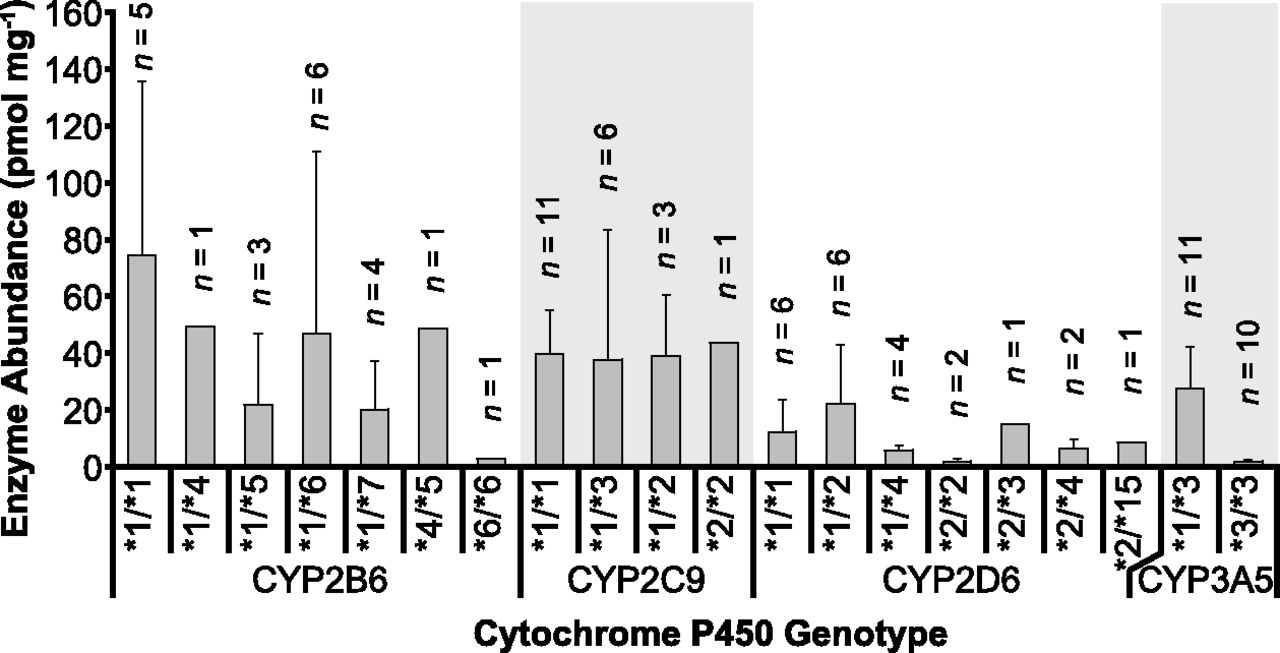
Mean abundance values of the different genotype variants of CYP2B6, CYP2C9, CYP2D6, and CYP3A5. Statistically significant difference was found between abundances of CYP3A5*1/*3 and CYP3A5*3/*3 (U-test, P < 0.0001) and between CYP2D6 variants (nonparametric ANOVA, P = 0.05). Error bars represent standard deviation values.
Relationship between Abundance of Enzymes and Age
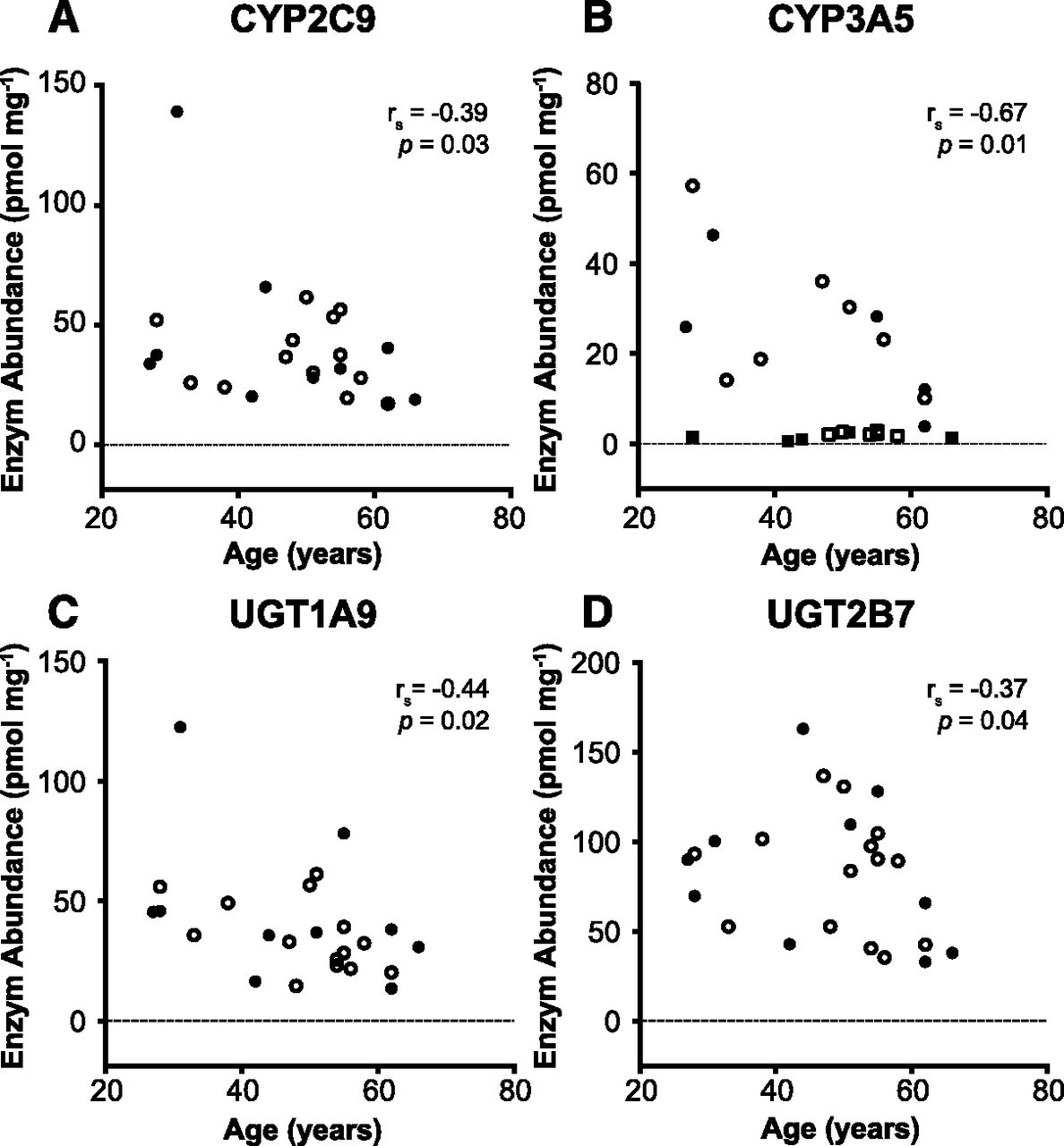
The expression of cytochrome P450 and UGT enzymes declined with age in general. However, statistically significant relationships were identified only in the case of CYP2C9 (rs = –0.39, P = 0.03, n = 23), CYP3A5*1/*3 (rs = –0.67, P = 0.01, n = 12), UGT1A3 (rs = –0.38, P = 0.04, n = 23), UGT1A9 (rs = –0.44, P = 0.02, n = 24), and UGT2B7 (rs = –0.37, P = 0.04, n = 24). A strong correlation was noted for the effects of age on CYP3A5 in individuals with *1/*3 genotype (Fig. 5). This was not the case for individuals with *3/*3 genotype. The effect of age on CYP3A5 would not have been recognized were the data not stratified by genotype (rs = –0.23, P = 0.14, n = 23).

Correlation between age of donors and abundance of CYP2C9 (A), CYP3A5 (B), UGT1A9 (C), and UGT2B7 (D). A declining expression trend can be seen with age in all cases. Age correlations are statistically significant (P < 0.05). Open symbols, male; closed symbols, female subjects. In B: circles, CYP3A5*1/*3; squares, CYP3A5*3/*3 variant.
Relationship between Abundance of Enzymes and Sex
No statistically significant differences between abundance data of cytochrome P450 and UGT enzymes in male and female subjects were seen.
Relationship between Abundance of Enzymes and Smoking and Alcohol Consumption
No significant differences of expression were observed between smokers and nonsmokers. In addition, no statistically significant differences were observed between abundances of enzymes in subjects who used alcohol and those who did not.
Relationship between Enzyme Abundance and Activity
Activity data were available for CYP1A2, CYP2B6, CYP2C9, CYP2D6, and CYP3A. There were strong and statistically significant abundance-activity correlations for CYP1A2 (rs = 0.80, P < 0.0001), CYP2B6 (rs = 0.82, P < 0.0001), CYP2D6 (rs = 0.65, P = 0.0006), and CYP3A4 (rs = 0.79, P < 0.0001). However, the relationship between the abundance of CYP2C9 and its activity was weak and of borderline significance (rs = 0.42, P = 0.05) (Fig. 6).

Correlation of enzyme activity and abundance of CYP1A2 (A), CYP2B6 (B), CYP2D6 (C), CYP3A4 (D), and CYP2C9 (E).
Discussion
Knowledge of the abundance of drug-metabolizing enzymes is essential for extrapolation of information on metabolic clearance (expressed per unit of enzyme) obtained from in vitro studies. Drug development requires optimization of pharmacokinetic properties, frequently based on prediction of in vivo behavior from in vitro measurements. The scaling factors required for IVIVE and the differences between the various in vitro systems are described elsewhere (Barter et al., 2007).
The present study quantified abundance of two families of enzymes involved in drug metabolism, P450s and UGTs, in the commonly used human liver microsomal in vitro system, using a QconCAT-based SRM assay (Russell et al., 2013) and established correlations between these enzymes within the set of samples studied. These correlations form an important element of population-based in silico prediction of absorption, distribution, metabolism, and excretion (ADME) (Rostami-Hodjegan and Tucker, 2007) and ignoring them may lead to erroneous assignment of variability (Barter et al., 2010; Cubitt et al., 2011). P450 and UGT enzymes were estimated to account for over 90% of metabolic elimination of the top 200 most prescribed drugs in 2002 (Wienkers and Heath, 2005). Although the current estimate might be different, it is unlikely that the importance of these enzymes has changed dramatically.
Several SRM assays have been applied to quantification of P450 and UGT enzymes (Ohtsuki et al., 2012; Sridar et al., 2013); however, this study is the first based on the QconCAT (Beynon et al., 2005) technique. For two of the enzymes (CYP3A4 and CYP3A5), we confirmed consistency with immunoquantification carried out previously. Interestingly, the immunoquantification measurements were systematically higher (1.5-fold on average) than the LC-MS assay. LC-MS is potentially susceptible to underestimation of protein concentration because of possible incomplete release of analytical peptide from the gel or inefficient proteolytic cleavage, but immunoquantification is susceptible to overestimation of protein abundance because of the possibility of cross-reactivity of primary antibodies with homologous enzymes (Fallon et al., 2008). In view of these possible sources of error, the agreement between the two sets of measurements is remarkable, and the fact that the discrepancies show a consistent pattern is especially encouraging. The LC-MS assay was shown to be linear over 1- to 32-fold dilution range, which is consistent with the dynamic range of P450 and UGT expression levels, and is precise, with coefficients of variation of abundance measurements not exceeding 18%.
We successfully quantified 13 out of the 15 P450 enzymes simultaneously. The other two enzymes had previously been quantified by immunoquantification methods. We speculate that further experimentation would yield quantification of these enzymes. CYP2E1 may require a larger QconCAT spike because of its high abundance in our samples while the low abundance CYP2C19 may require a larger sample concentration. We also quantified 8 out of 10 UGTs successfully. The remaining two UGTs are reported to have low abundance and only a single, long, potentially post-translationally modified peptide was available for quantification of each of them. An attempt to quantify UGT1A11 was recently published (Fallon et al., 2013).
Quantification of enzyme abundance in these samples revealed large interindividual variations in expression for P450 enzymes (5- to 500-fold). The variation was much less for expression of UGT enzymes (5- to 20-fold). The expected variability related to analytical error [i.e., fold difference between the 5th and 95th centiles; (1 + 2 CV)/(1 – 2 CV)] was only 2-fold.
Virtual patients in simulated drug trials must exhibit realistic combinations of enzyme expression. The most notable correlations found in the literature include a strong relationship between CYP3A4 and CYP3A5*1/*3 (Barter et al., 2010). This correlation was confirmed in the present study, while such strong relationship was absent for other CYP3A5 variants. Other strong and statistically significant correlations reported in the literature include those between the pairs: CYP3A4/CYP2C8 (McSorley and Daly, 2000; Naraharisetti et al., 2010), CYP2C8/CYP2C9 (Edwards et al., 1998), and CYP3A4/CYP2B6 (Mimura et al., 1993; Totah et al., 2008); all of these were also confirmed in our set of samples. These results, together with the mRNA analysis of Wortham et al. (2007), support the reported common genetic regulation mechanisms (Jover et al., 2009), which involve nuclear receptors.
The wide range of enzymes quantified in the present study allowed novel correlations to be demonstrated in the same subjects (n = 24). The only correlation between expression of UGT enzymes reported previously is a weak interaction between UGT1A1 and UGT1A6 (correlation coefficient = 0.33, n = 10) (Fallon et al., 2008). Our results are consistent (rs = 0.42, P = 0.02, n = 23). We have also found, however, strong and significant correlations between the pairs: UGT1A1/UGT2B4, UGT1A4/UGT2B4, UGT1A6/UGT1A9, UGT1A9/UGT2B4, UGT2B4/UGT2B15, and UGT2B7/UGT2B15. These data are summarized in Table 5.
The QconCAT methodology is especially powerful for the measurement of correlations between abundances of various proteins. For example, cross-correlation analysis between P450 and UGT expression at the protein level has not been described previously, although Wortham et al. (2007) demonstrated possible correlations of expression between UGT1A1 and some P450 enzymes at the mRNA level. In the present study, some significant relationships are described for the first time: CYP1A2/UGT2B4, CYP1A2/UGT2B7, CYP2C9/UGT1A4, CYP2C9/UGT2B4, and CYP2C9/UGT2B7. Many xenobiotics are first metabolized by cytochrome P450 enzymes and then glucuronidated and these correlations are certainly mechanistically reasonable; nevertheless, it would be reassuring to see them repeated in larger numbers of samples.
Genotypes for some P450 enzymes were provided by the supplier, enabling correlation of enzyme abundance with genotype. Samples with CYP3A5*1/*3 genotype showed enzyme expression 14-fold higher than samples with CYP3A5*3/*3 genotype (P < 0.0001), as supported by the literature (Kuehl et al., 2001; Lin et al., 2002; Westlind-Johnsson et al., 2003). Although statistically significant differences in expression were previously reported between different genotypes of CYP2D6 (Zanger et al., 2001), these differences were not confirmed by the present study, most likely due to small sample size for individual genotypes.
Correlations of age, sex, smoking, and alcohol consumption with P450 and UGT abundances were investigated. Cytochrome P450 and UGT enzyme abundance declined consistently with age, but the correlation was statistically significant only in the cases of CYP2C9, CYP3A5*1/*3, UGT1A3, UGT1A9, and UGT2B7 (P < 0.05). This study included only adult subjects and therefore the established relationship of enzyme expression with age until adulthood (ontogeny) was not observed (Al Omari and Murry, 2007; Court, 2010). In adult subjects, age was reported to have little effect on the abundance and activity of cytochrome P450 (Hunt et al., 1990, 1992; Schmucker et al., 1990; Shimada et al., 1994) and on the activity of UGT enzymes (Herd et al., 1991; Court, 2010). Analysis of reported metabolic ratios indicated a decline in metabolic activity that parallels the decline in renal function (Rostami-Hodjegan et al., 1999); however, liver size shrinkage as body size decreases with age (Polasek et al., 2013) may be a contributing factor.
There were little differences in enzyme expression between male and female subjects. Wolbold et al. (2003) reported higher CYP3A4 expression in female than male subjects. Gender had little effect on the levels of UGT enzymes in the analyzed samples, in line with literature on UGT activity (Court, 2010). Effects of gender differences on drug metabolism have previously been investigated (Chetty et al., 2012).
CYP1A2 is known to be upregulated by smoking (Kalow and Tang, 1991) and clearance of CYP1A2 probes correlates with the level of smoking. Failure to detect significant differential expression of CYP1A2 may be due to small sample size and absence of information on the level of smoking by the subjects. One sample from a subject who was assigned by the supplier as a smoker had to be reassigned to nonsmoker status in this study as the detailed description indicated that she had stopped smoking 11 years before donation. Such detailed information was unavailable for other donors. Higher activity of UGT enzymes in smokers has previously been reported (Court, 2010); however, as with P450s, we were unable to detect statistically significant differences in expression.
Alcohol consumption did not have a significant effect on measured cytochrome P450 or UGT expression. Induction of CYP2E1, which metabolizes alcohol, has previously been observed with chronic alcohol consumption (Lieber, 1988); however, CYP2E1 was not quantified in this study. Poor characterization of drinking habits in these subjects made identifying groups of heavy drinkers within the subjects impossible. Application of this assay to sample sets better characterized with respect to smoking, alcohol consumption, and medication may reveal more subtle correlations with these environmental factors.
Activity of cytochrome P450 enzymes and their abundance correlated strongly in the cases of CYP1A2, 2B6, 2D6, and 3A4, but not for CYP2C9. This could reflect inaccurate activity measures for CYP2C9, which were determined with a less specific probe (diclofenac) as opposed to the more selective S-warfarin (Tucker et al., 2001). Alternatively, abundance may not reflect the activity when various CYP2C9 genotypes are assessed together. In addition, differences in expression of cytochrome b5 and NADPH P450 reductase in different individuals have been reported to lead to disparities in CYP2C9 enzyme activity (Crewe et al., 2011). These were not known for individual samples under study.
Despite relatively small sample size used here compared with previous reports that used immunoblotting to assess abundances of P450 and UGT enzymes, this report, as a proof of principle, demonstrated correlations reported previously and uncovered novel correlations between expression levels of different enzymes. Larger sample numbers are expected to yield more comprehensive correlation networks not just between enzymes but also with drug transporters. The QconCAT method, despite its limitations, represents an improvement over established methodology for simultaneous quantification of liver enzymes with cost implications with respect to providing more sustainable standards.
Acknowledgments
The authors thank Pfizer (USA) for supplying the liver microsomal samples, Maurice Dickens and Michael Zientek for supplying enzyme activity data alongside the liver samples, and Eleanor Savill for assisting with preparation of the manuscript.
Authorship Contributions
Participated in research design: Achour, Barber, Rostami-Hodjegan.
Conducted experiments: Achour, Russell.
Contributed new reagents or analytic tools: Achour, Barber, Rostami-Hodjegan.
Performed data analysis: Achour, Barber, Rostami-Hodjegan.
Wrote or contributed to the writing of the manuscript: Achour, Russell, Barber, Rostami-Hodjegan.
Footnotes
- Received October 31, 2013.
- Accepted January 9, 2014.
The authors thank Khaled Hassan Al-Ghamdi Est. for funding B. Achour, the Manchester Pharmacy School for funding M. Russell, and Simcyp Limited (a Certara company) for financial support toward the project.
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- HLM
- human liver microsomes
- IVIVE
- in vitro-in vivo extrapolation
- LC-MS
- liquid chromatography in conjunction with mass spectrometry
- MetCAT
- metabolism enzymes quantification concatemer
- MS/MS
- tandem mass spectrometry
- NNOP
- nonnaturally occurring peptide
- P450
- cytochrome P450
- QconCAT
- quantification concatemer
- SRM
- selected reaction monitoring
- UGT
- uridine 5′-diphospho-glucuronosyltransferase
- Copyright © 2014 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}